

Elongation during segmentation shows axial variability, low mitotic rates, and synchronized cell cycle domains in the crustacean, Thamnocephalus platyurus
- PMID: 31988708
- PMCID: PMC6969478
- DOI: 10.1186/s13227-020-0147-0
Elongation during segmentation shows axial variability, low mitotic rates, and synchronized cell cycle domains in the crustacean, Thamnocephalus platyurus
Abstract
Background: Segmentation in arthropods typically occurs by sequential addition of segments from a posterior growth zone. However, the amount of tissue required for growth and the cell behaviors producing posterior elongation are sparsely documented.
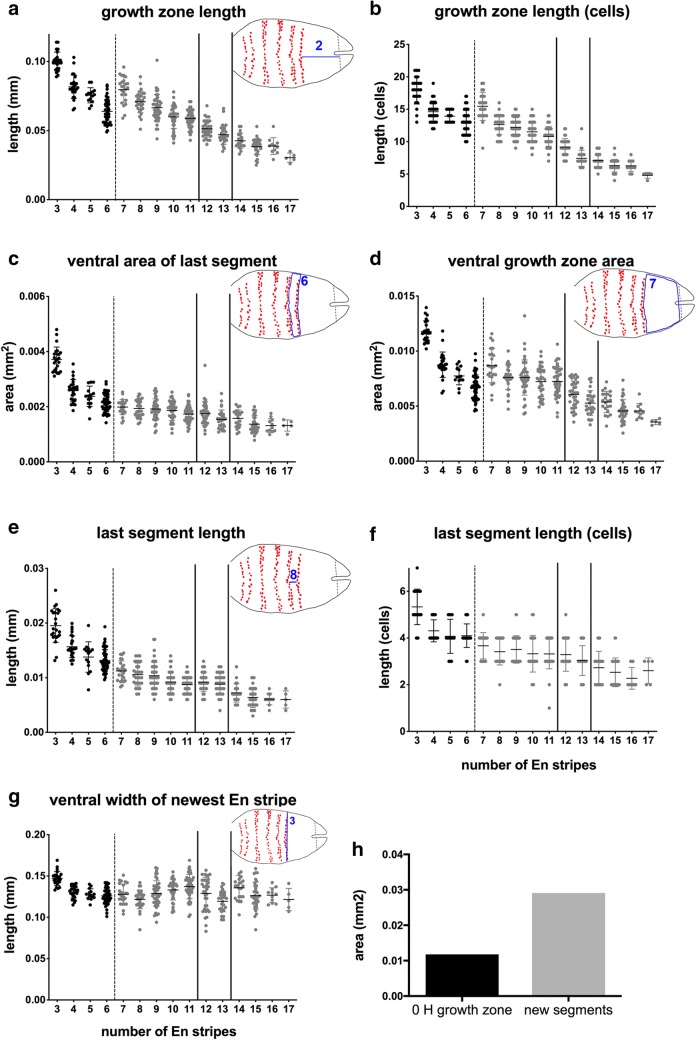
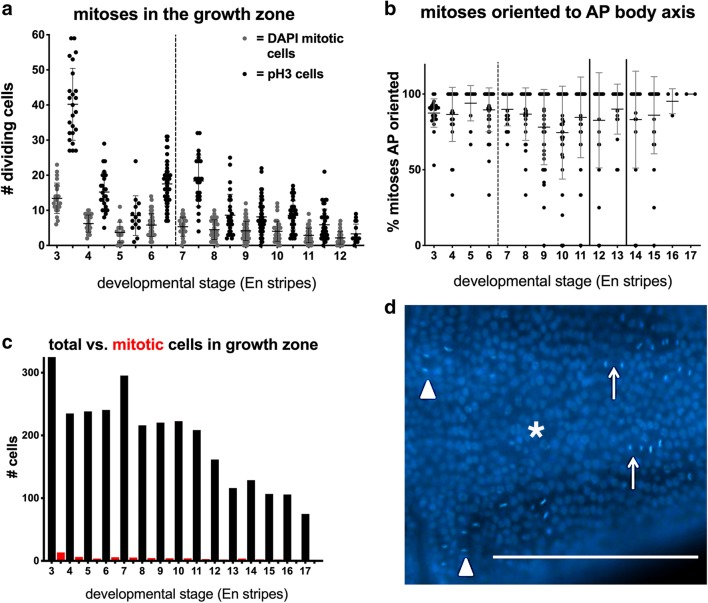
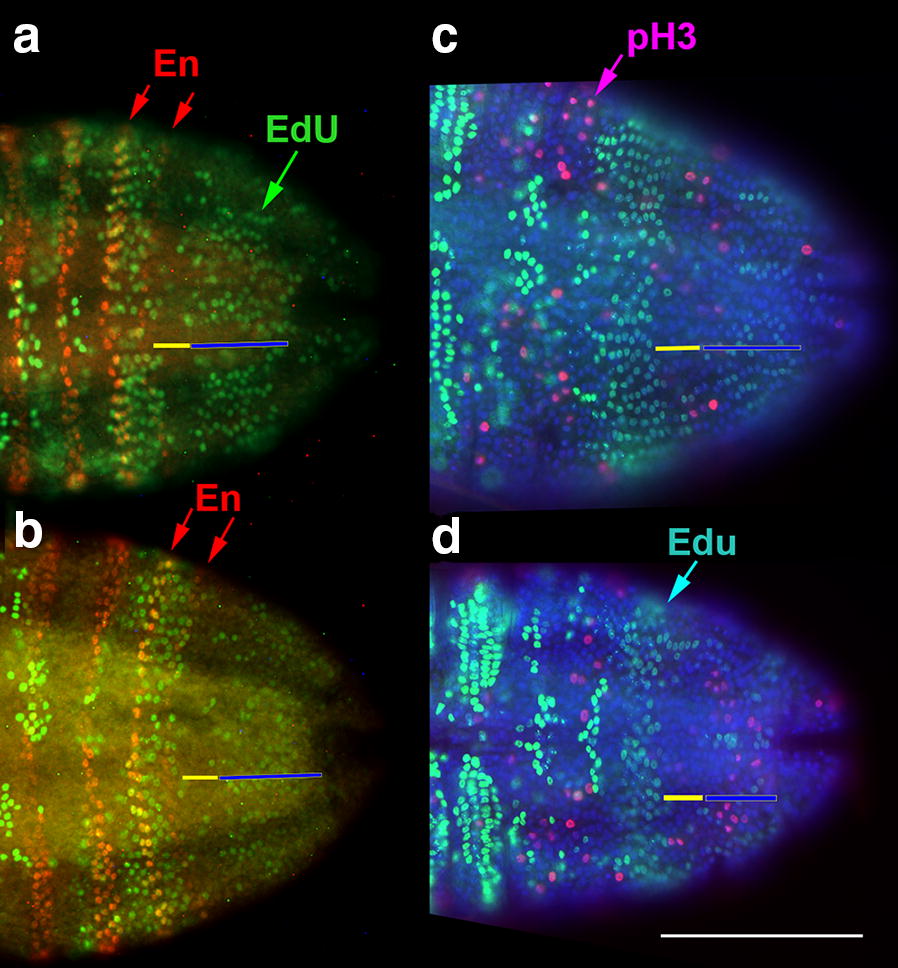
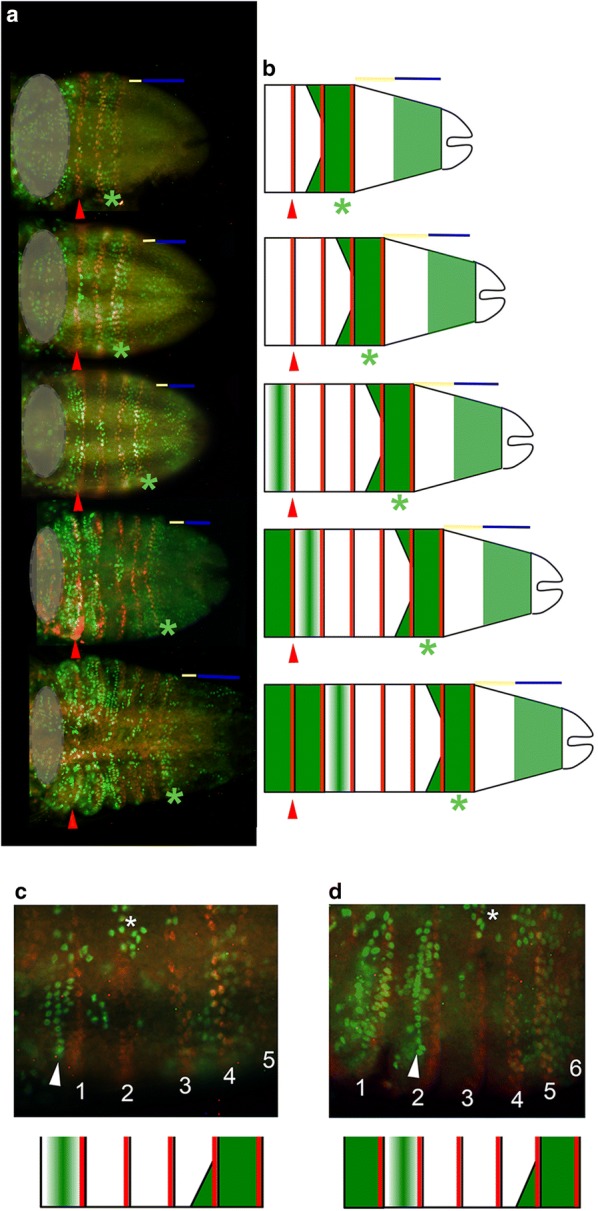
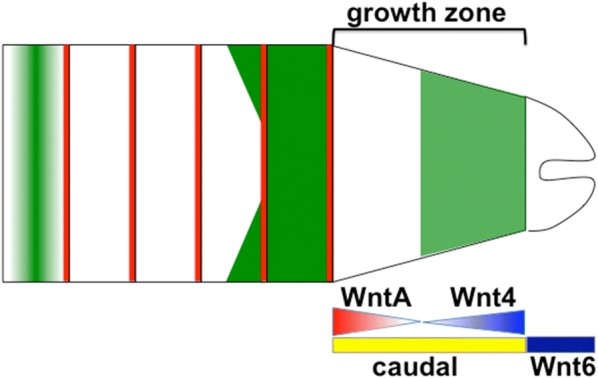
Results: Using precisely staged larvae of the crustacean, Thamnocephalus platyurus, we systematically examine cell division patterns and morphometric changes associated with posterior elongation during segmentation. We show that cell division occurs during normal elongation but that cells in the growth zone need only divide ~ 1.5 times to meet growth estimates; correspondingly, direct measures of cell division in the growth zone are low. Morphometric measurements of the growth zone and of newly formed segments suggest tagma-specific features of segment generation. Using methods for detecting two different phases in the cell cycle, we show distinct domains of synchronized cells in the posterior trunk. Borders of cell cycle domains correlate with domains of segmental gene expression, suggesting an intimate link between segment generation and cell cycle regulation.
Conclusions: Emerging measures of cellular dynamics underlying posterior elongation already show a number of intriguing characteristics that may be widespread among sequentially segmenting arthropods and are likely a source of evolutionary variability. These characteristics include: the low rates of posterior mitosis, the apparently tight regulation of cell cycle at the growth zone/new segment border, and a correlation between changes in elongation and tagma boundaries.
Keywords: Arthropod; EdU; Growth zone; Mitosis; Segmentation; Wnt.
© The Author(s) 2020.
Conflict of interest statement
Competing interestsThe authors declare that they have no competing interests.
Figures




References
-
- Lawrence PA. The making of a fly: the genetics of animal design. Oxford: Blackwell Scientific Publications; 1992.
-
- Chipman AD. Thoughts and speculations on the ancestral arthropod segmentation pathway. In: Fusco G, editor. Evolving pathways: key themes in evolutionary developmental biology. Cambridge: Cambridge University Press; 2008. pp. 342–356.
-
- Scholtz G. Teloblasts in decapod embryos: an embryonic character reveals the monophyletic origin of freshwater crayishes (Crustacea, Decapoda) Zool Anz. 1993;230:45–54.
-
- Snodgrass RE. Evolution of the annelida, onychophora, and arthropoda. Smithson Misc Collect. 1938;97:1–159.
-
- Anderson DT. Larval development and segment formation in the branchiopod crustaceans Limnadia stanleyana King (Conchostraca) and Artemia salina (L.) (Anostraca). Aust. J Zool. 1967;15:47–91.
LinkOut - more resources
Full Text Sources
