

Autophagy of an Amyloid-like Translational Repressor Regulates Meiotic Exit
- PMID: 31991104
- PMCID: PMC7138260
- DOI: 10.1016/j.devcel.2019.12.017
Autophagy of an Amyloid-like Translational Repressor Regulates Meiotic Exit
Abstract
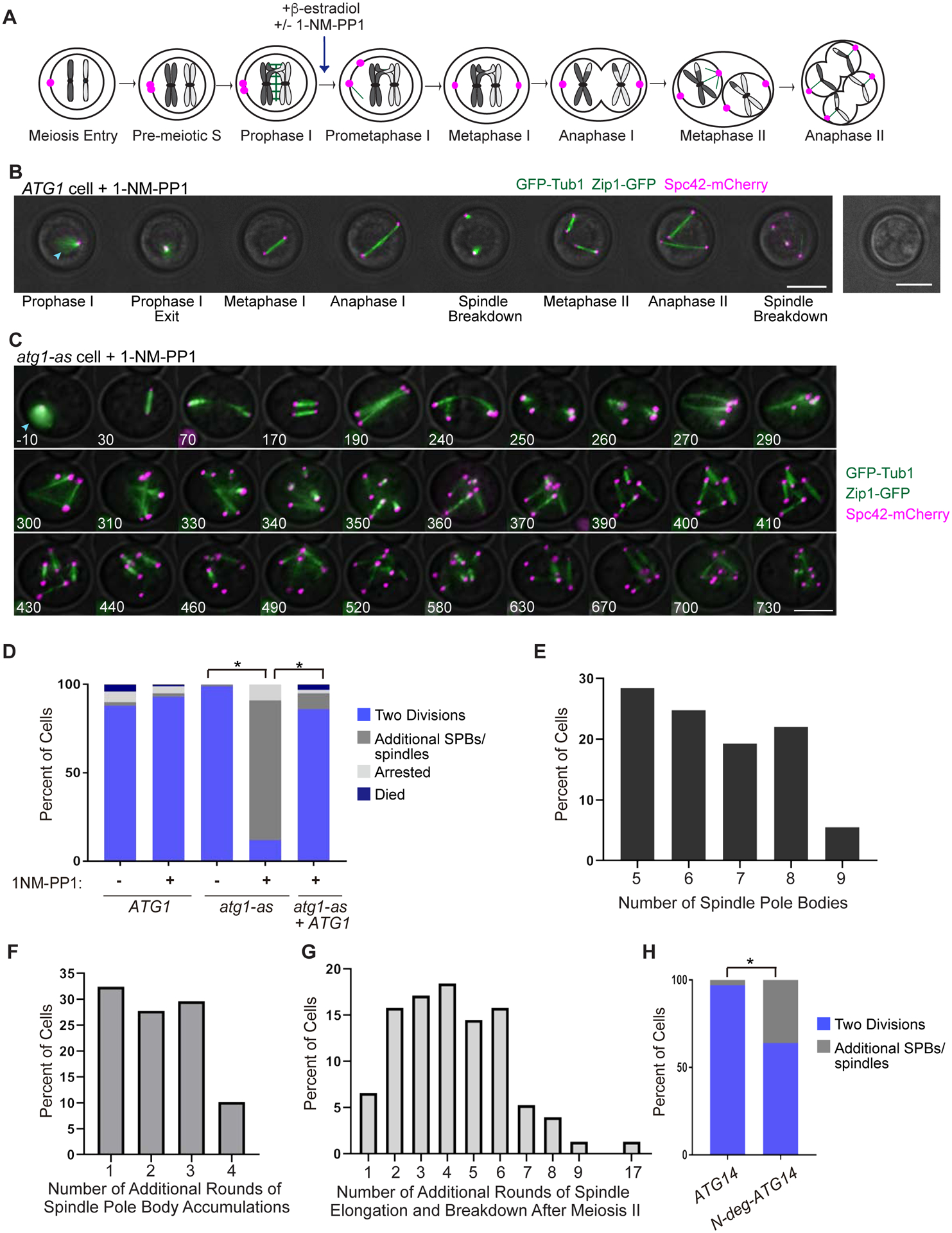
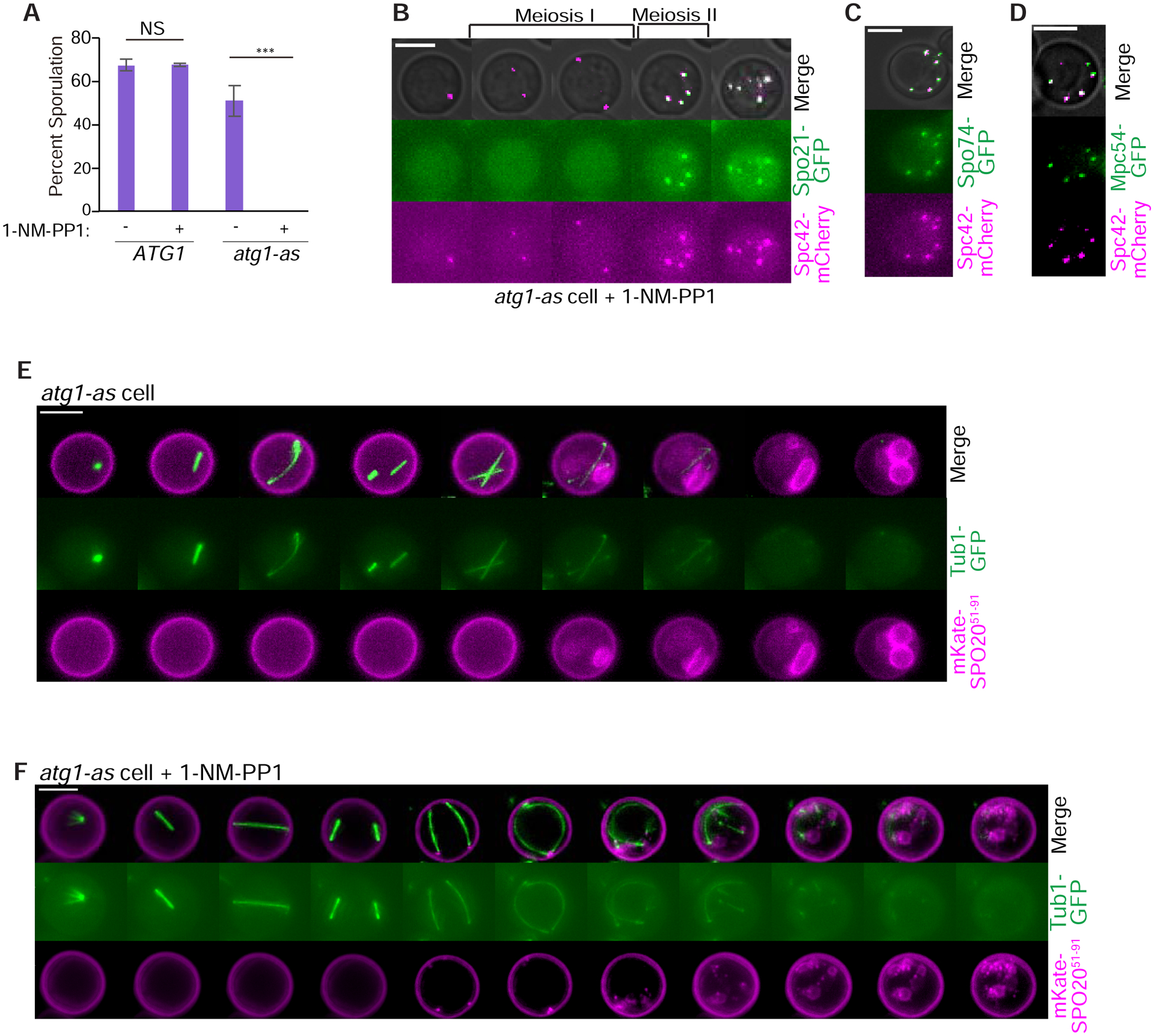
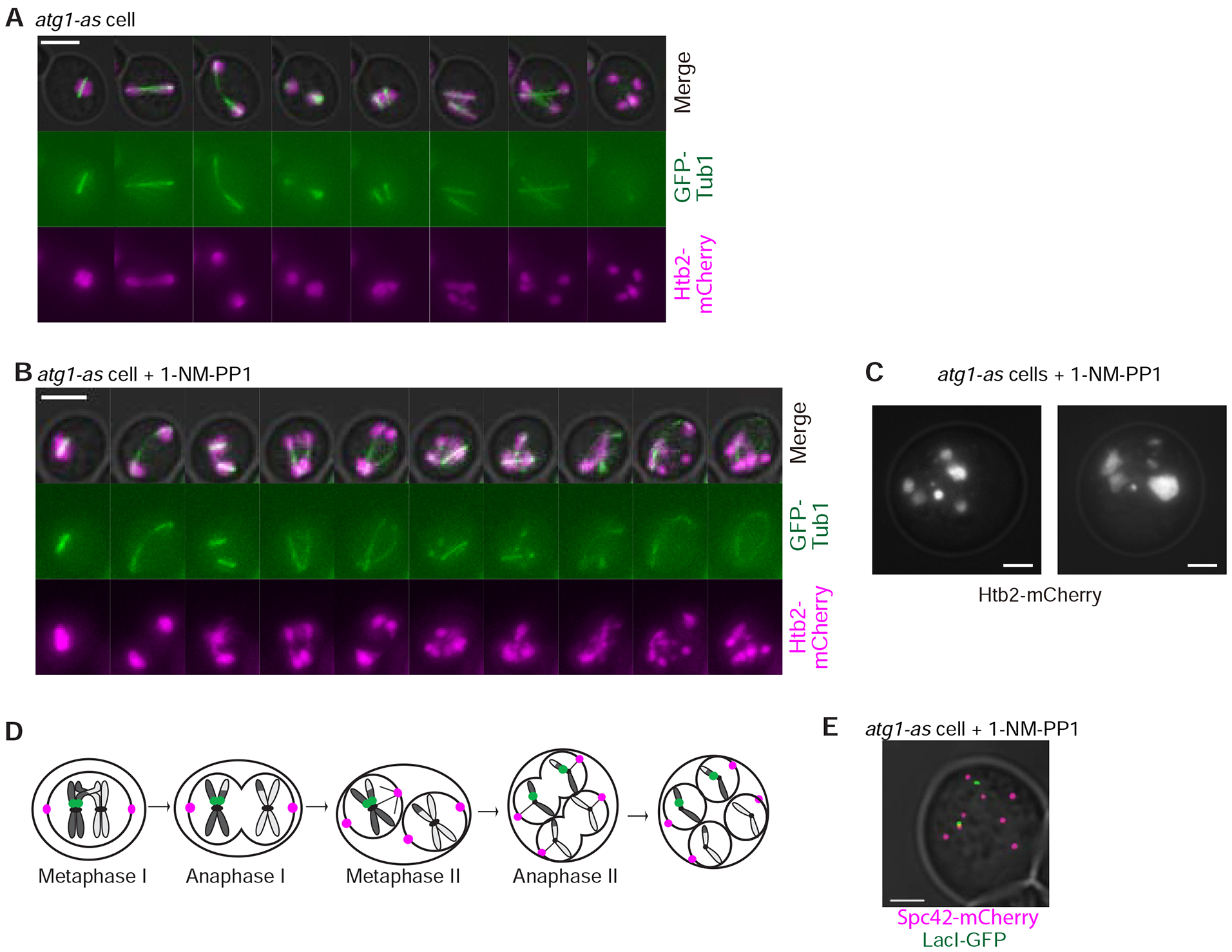
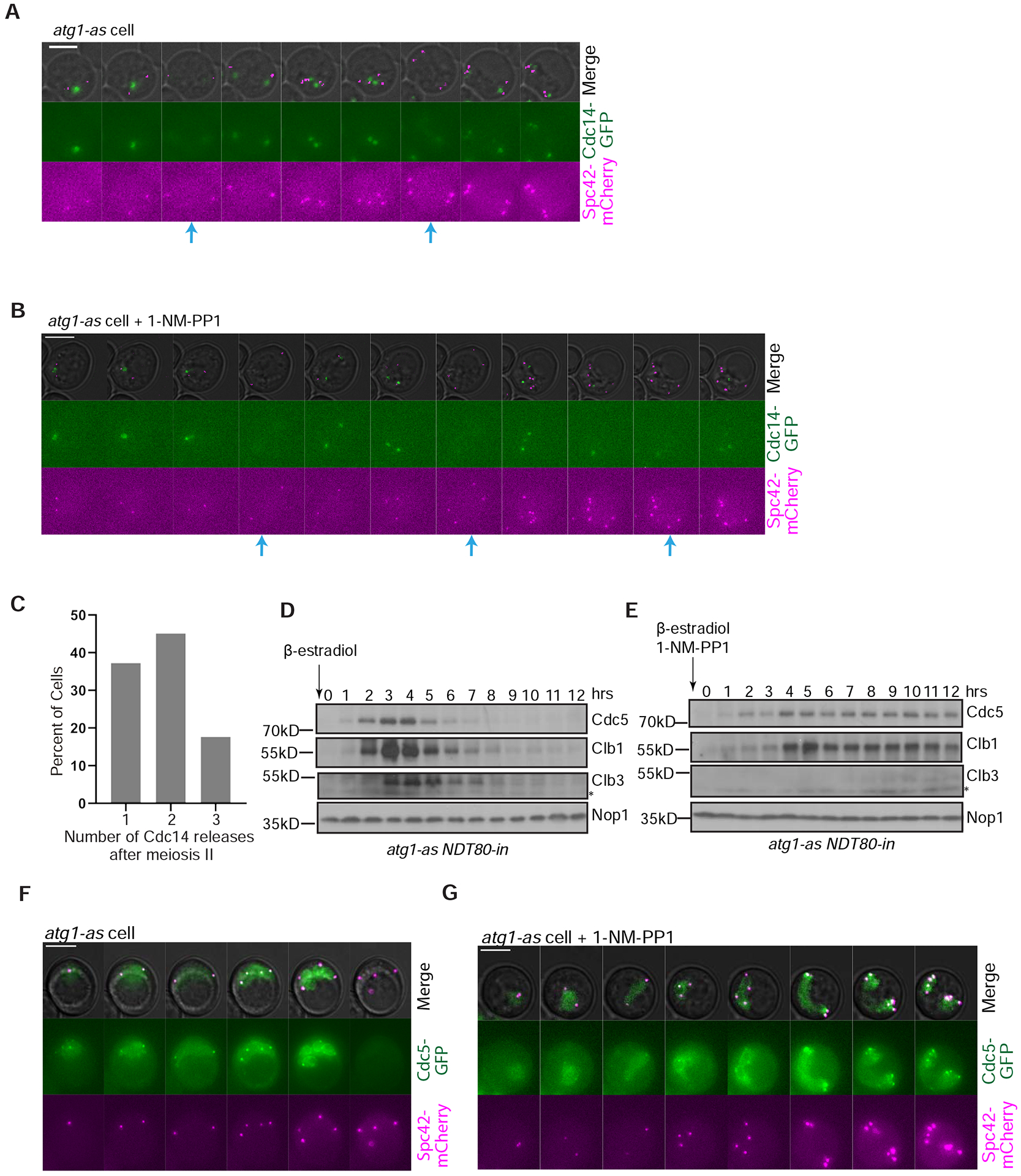
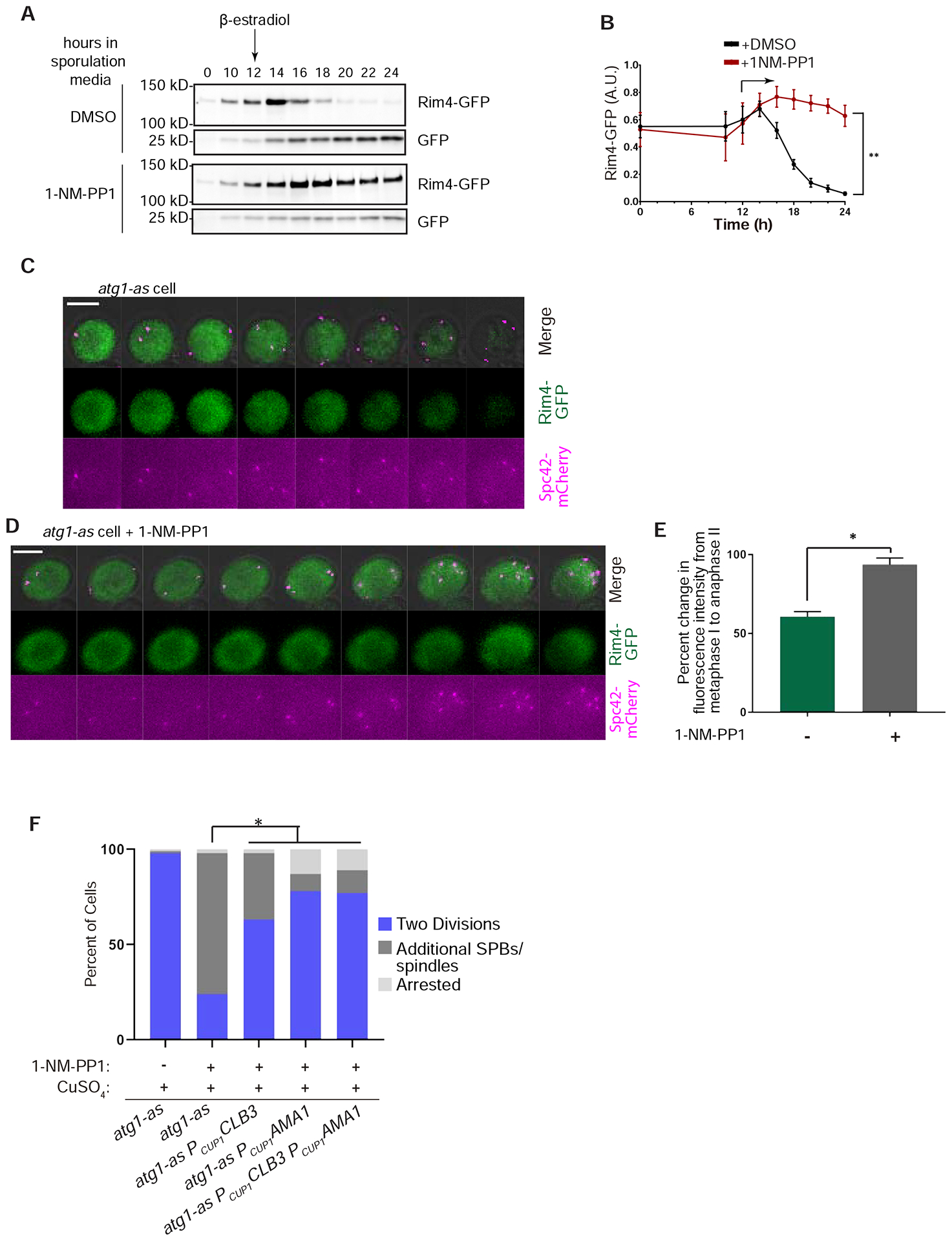
We explored the potential for autophagy to regulate budding yeast meiosis. Following pre-meiotic DNA replication, we blocked autophagy by chemical inhibition of Atg1 kinase or engineered degradation of Atg14 and observed homologous chromosome segregation followed by sister chromatid separation; cells then underwent additional rounds of spindle formation and disassembly without DNA re-replication, leading to aberrant chromosome segregation. Analysis of cell-cycle regulators revealed that autophagy inhibition prevents meiosis II-specific expression of Clb3 and leads to the aberrant persistence of Clb1 and Cdc5, two substrates of a meiotic ubiquitin ligase activated by Ama1. Lastly, we found that during meiosis II, autophagy degrades Rim4, an amyloid-like translational repressor whose timed clearance regulates protein production from its mRNA targets, which include CLB3 and AMA1. Strikingly, engineered Clb3 or Ama1 production restored meiotic termination in the absence of autophagy. Thus, autophagy destroys a master regulator of meiotic gene expression to enable irreversible meiotic exit.
Keywords: APC/C; Ama1; Atg1; Atg14; Clb3; Rim4; autophagy; meiosis; sporulation; translational repression.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Arguello-Miranda O, Zagoriy I, Mengoli V, Rojas J, Jonak K, Oz T, Graf P, and Zachariae W (2017). Casein Kinase 1 Coordinates Cohesin Cleavage, Gametogenesis, and Exit from M Phase in Meiosis II. Developmental cell 40, 37–52. - PubMed
-
- Blethrow J, Zhang C, Shokat KM, and Weiss EL (2004). Design and use of analog-sensitive protein kinases. Curr Protoc Mol Biol Chapter 18, Unit 18 11. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
