

Large deletions in immunoglobulin genes are associated with a sustained absence of DNA Polymerase η
- PMID: 31992747
- PMCID: PMC6987143
- DOI: 10.1038/s41598-020-58180-7
Large deletions in immunoglobulin genes are associated with a sustained absence of DNA Polymerase η
Abstract
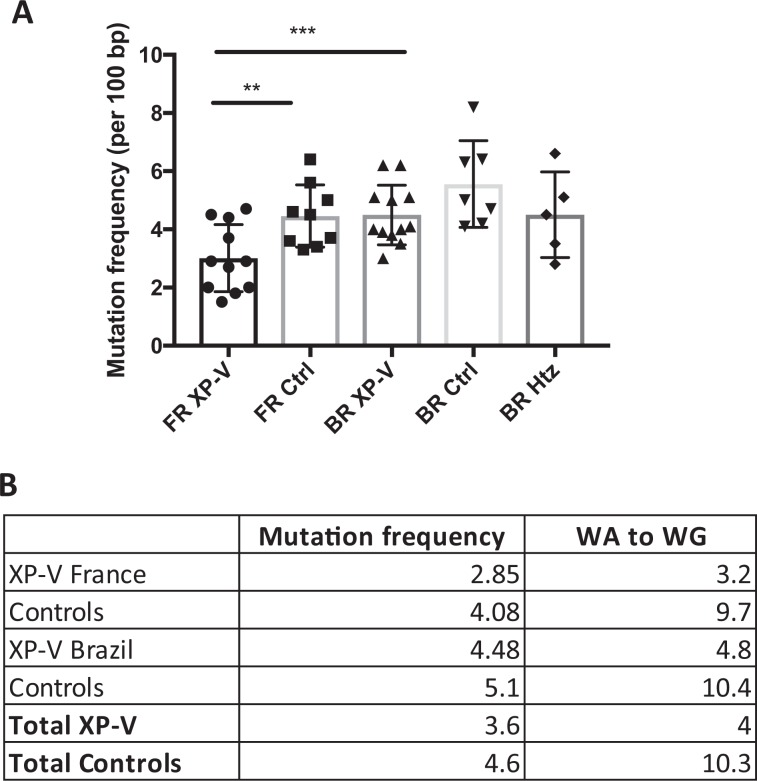
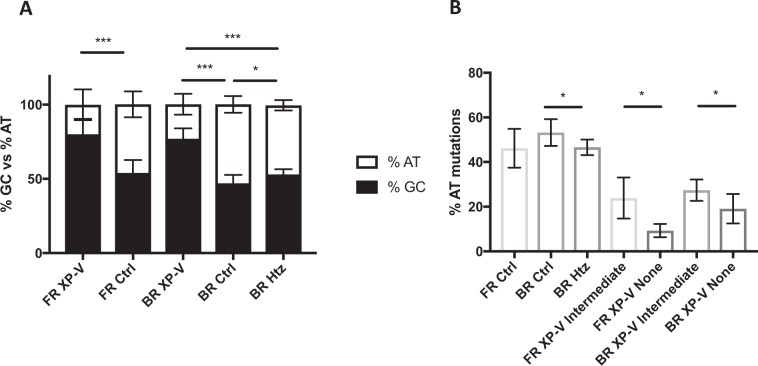
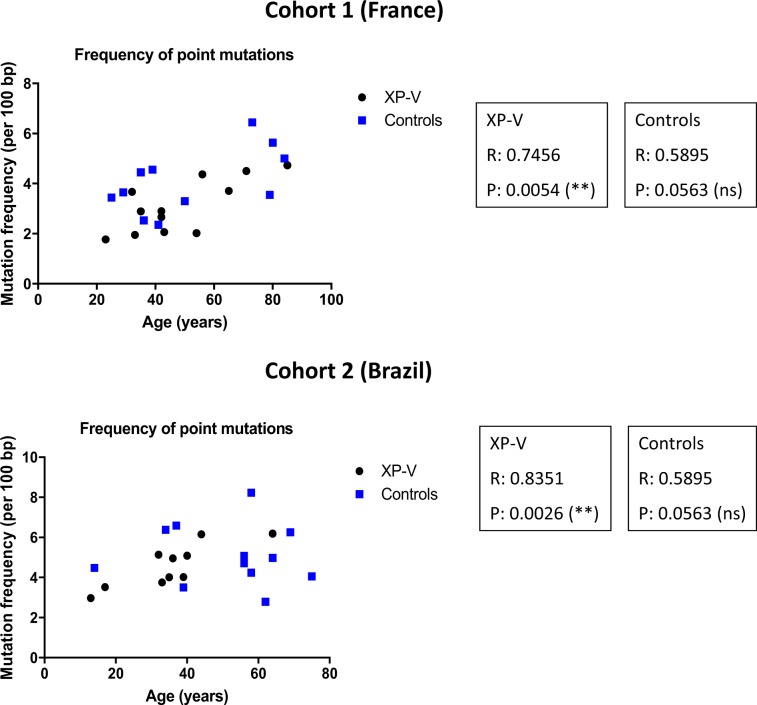
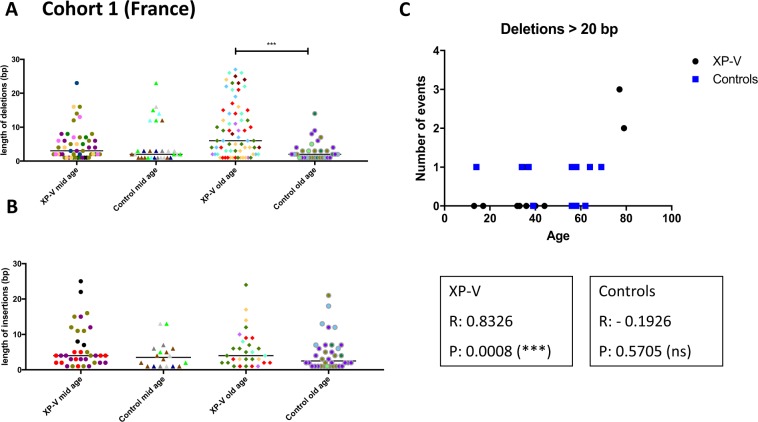
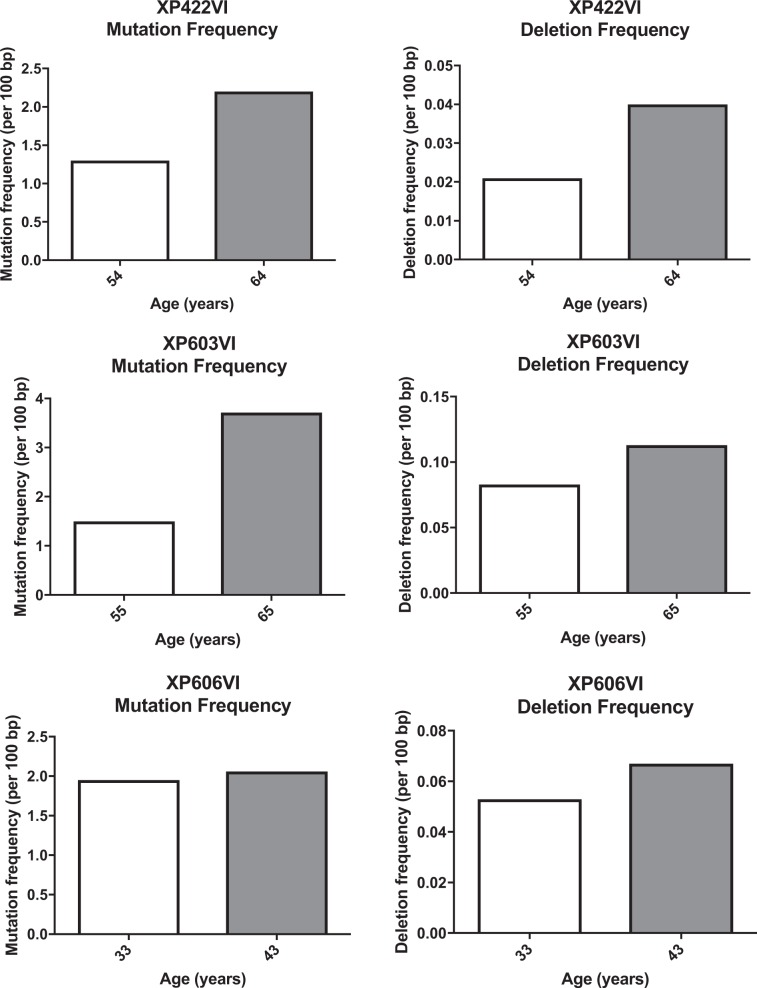
Somatic hypermutation of immunoglobulin genes is a highly mutagenic process that is B cell-specific and occurs during antigen-driven responses leading to antigen specificity and antibody affinity maturation. Mutations at the Ig locus are initiated by Activation-Induced cytidine Deaminase and are equally distributed at G/C and A/T bases. This requires the establishment of error-prone repair pathways involving the activity of several low fidelity DNA polymerases. In the physiological context, the G/C base pair mutations involve multiple error-prone DNA polymerases, while the generation of mutations at A/T base pairs depends exclusively on the activity of DNA polymerase η. Using two large cohorts of individuals with xeroderma pigmentosum variant (XP-V), we report that the pattern of mutations at Ig genes becomes highly enriched with large deletions. This observation is more striking for patients older than 50 years. We propose that the absence of Pol η allows the recruitment of other DNA polymerases that profoundly affect the Ig genomic landscape.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Miscellaneous
