

AMPA receptor deletion in developing MGE-derived hippocampal interneurons causes a redistribution of excitatory synapses and attenuates postnatal network oscillatory activity
- PMID: 31992779
- PMCID: PMC6987165
- DOI: 10.1038/s41598-020-58068-6
AMPA receptor deletion in developing MGE-derived hippocampal interneurons causes a redistribution of excitatory synapses and attenuates postnatal network oscillatory activity
Abstract
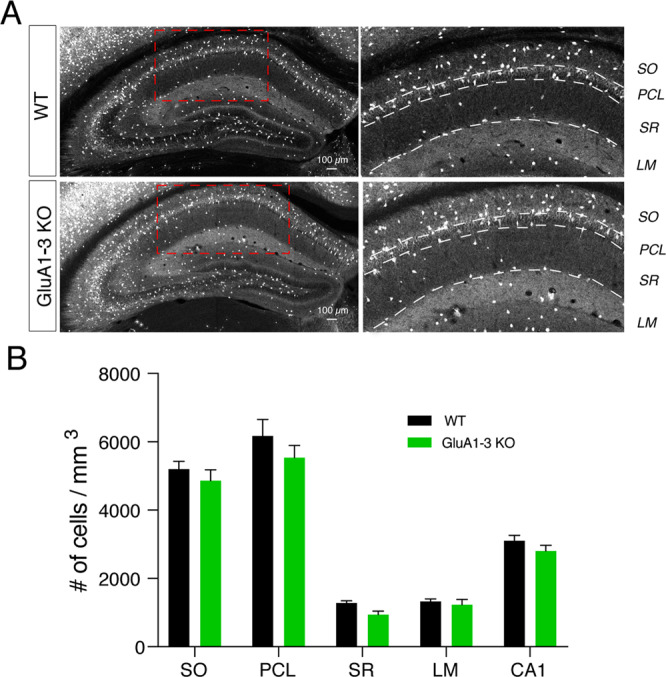
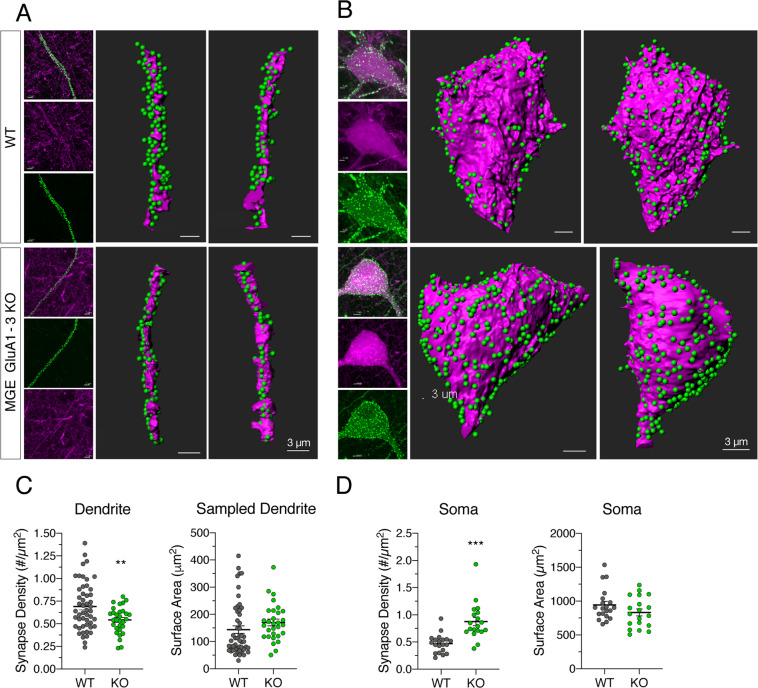
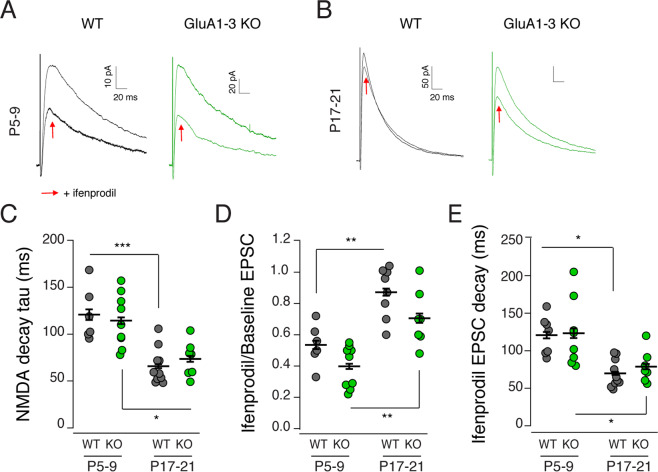
Inhibitory interneurons derived from the medial ganglionic eminence represent the largest cohort of GABAergic neurons in the hippocampus. In the CA1 hippocampus excitatory synapses onto these cells comprise GluA2-lacking, calcium-permeable AMPARs. Although synaptic transmission is not established until early in their postnatal life, AMPARs are expressed early in development, however their role is enigmatic. Using the Nkx2.1-cre mouse line we genetically deleted GluA1, GluA2, GluA3 selectively from MGE derived interneurons early in development. We observed that the number of MGE-derived interneurons was preserved in mature hippocampus despite early elimination of AMPARs, which resulted in >90% decrease in spontaneous excitatory synaptic activity. Of particular interest, excitatory synaptic sites were shifted from dendritic to somatic locations while maintaining a normal NMDAR content. The developmental switch of NMDARs from GluN2B-containing early in development to GluN2A-containing on maturation was similarly unperturbed despite the loss of AMPARs. Early network giant depolarizing potential oscillatory activity was compromised in early postnatal days as was both feedforward and feedback inhibition onto pyramidal neurons underscoring the importance of glutamatergic drive onto MGE-derived interneurons for hippocampal circuit function.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
