

Linear Ubiquitin Chains: Cellular Functions and Strategies for Detection and Quantification
- PMID: 31998699
- PMCID: PMC6966713
- DOI: 10.3389/fchem.2019.00915
Linear Ubiquitin Chains: Cellular Functions and Strategies for Detection and Quantification
Abstract
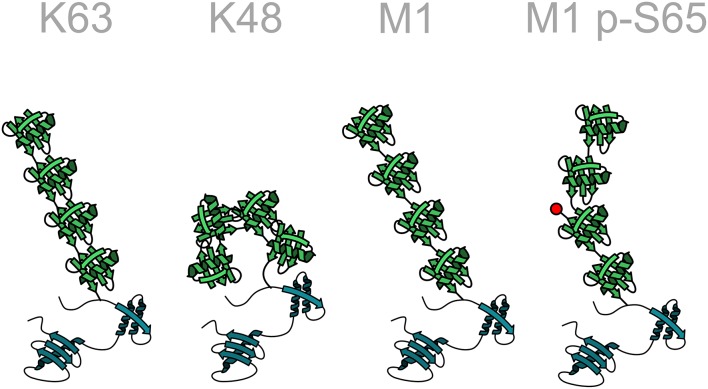
Ubiquitination of proteins is a sophisticated post-translational modification implicated in the regulation of an ever-growing abundance of cellular processes. Recent insights into different layers of complexity have shaped the concept of the ubiquitin code. Key players in determining this code are the number of ubiquitin moieties attached to a substrate, the architecture of polyubiquitin chains, and post-translational modifications of ubiquitin itself. Ubiquitination can induce conformational changes of substrates and alter their interactive profile, resulting in the formation of signaling complexes. Here we focus on a distinct type of ubiquitination that is characterized by an inter-ubiquitin linkage through the N-terminal methionine, called M1-linked or linear ubiquitination. Formation, recognition, and disassembly of linear ubiquitin chains are highly specific processes that are implicated in immune signaling, cell death regulation and protein quality control. Consistent with their role in influencing signaling events, linear ubiquitin chains are formed in a transient and spatially regulated manner, making their detection and quantification challenging.
Keywords: HOIL; HOIP; LUBAC; OTULIN; PRM; SHARPIN; SRM; ubiquitin.
Copyright © 2020 Dittmar and Winklhofer.
Figures








References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources