

Dissecting the Extracellular Complexity of Neuromuscular Junction Organizers
- PMID: 31998752
- PMCID: PMC6966886
- DOI: 10.3389/fmolb.2019.00156
Dissecting the Extracellular Complexity of Neuromuscular Junction Organizers
Abstract
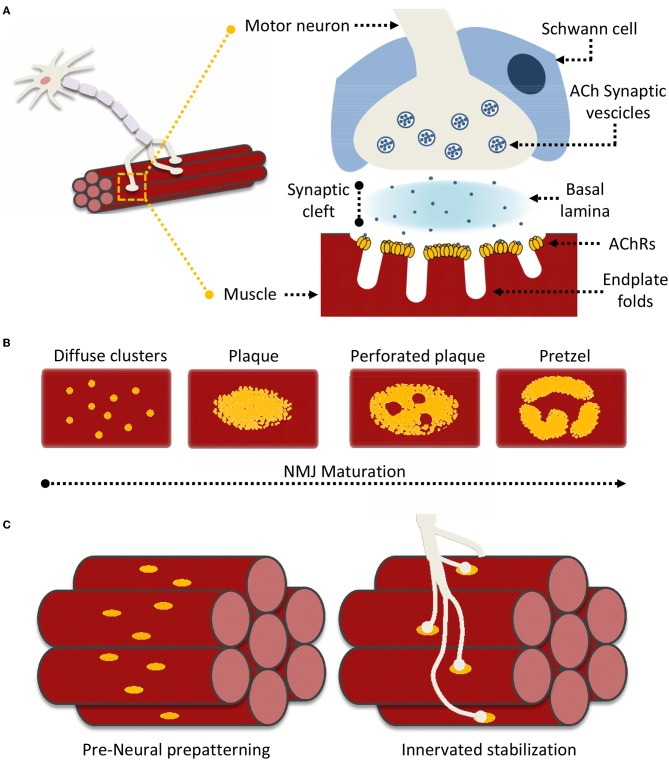
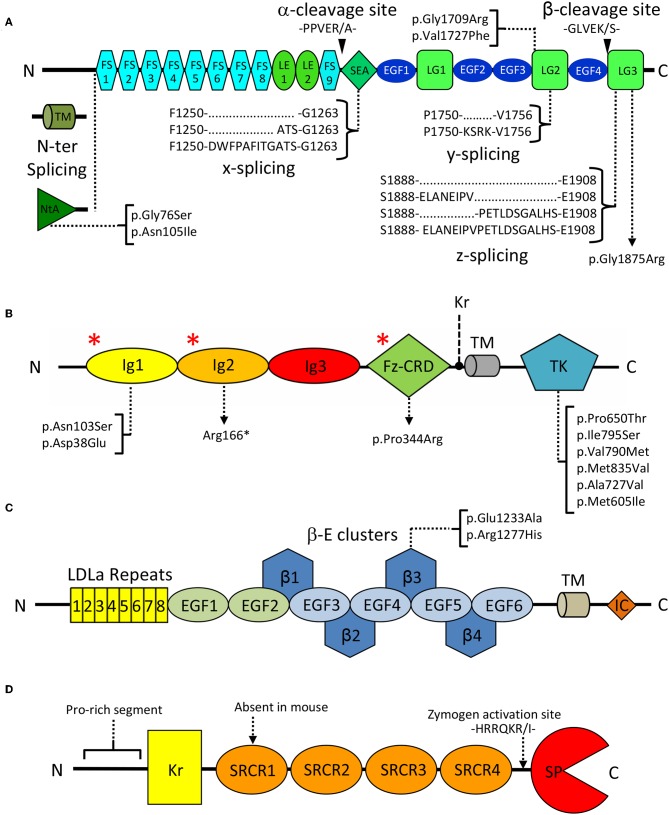
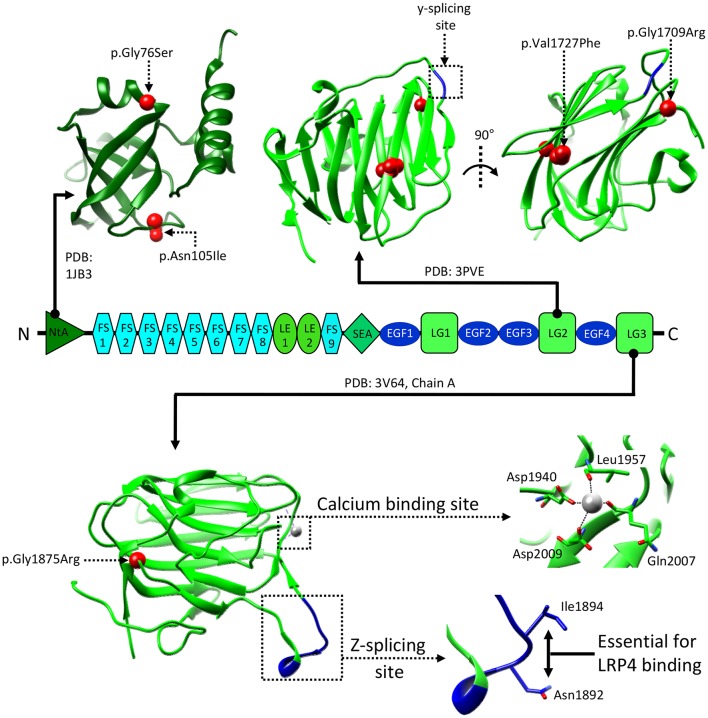
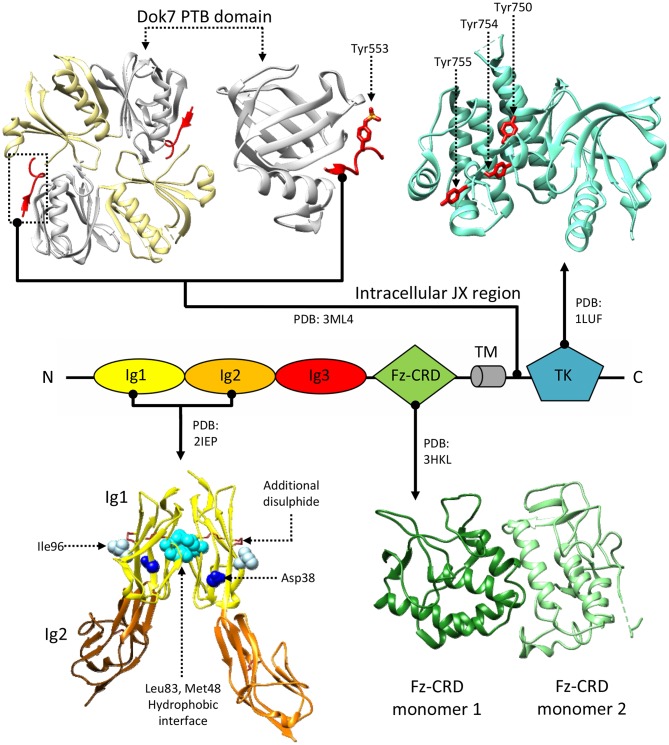
Synapse formation is a very elaborate process dependent upon accurate coordination of pre and post-synaptic specialization, requiring multiple steps and a variety of receptors and signaling molecules. Due to its relative structural simplicity and the ease in manipulation and observation, the neuromuscular synapse or neuromuscular junction (NMJ)-the connection between motor neurons and skeletal muscle-represents the archetype junction system for studying synapse formation and conservation. This junction is essential for survival, as it controls our ability to move and breath. NMJ formation requires coordinated interactions between motor neurons and muscle fibers, which ultimately result in the formation of a highly specialized post-synaptic architecture and a highly differentiated nerve terminal. Furthermore, to ensure a fast and reliable synaptic transmission following neurotransmitter release, ligand-gated channels (acetylcholine receptors, AChRs) are clustered on the post-synaptic muscle cell at high concentrations in sites opposite the presynaptic active zone, supporting a direct role for nerves in the organization of the post-synaptic membrane architecture. This organized clustering process, essential for NMJ formation and for life, relies on key signaling molecules and receptors and is regulated by soluble extracellular molecules localized within the synaptic cleft. Notably, several mutations as well as auto-antibodies against components of these signaling complexes have been related to neuromuscular disorders. The recent years have witnessed strong progress in the understanding of molecular identities, architectures, and functions of NMJ macromolecules. Among these, prominent roles have been proposed for neural variants of the proteoglycan agrin, its receptor at NMJs composed of the lipoprotein receptor-related protein 4 (LRP4) and the muscle-specific kinase (MuSK), as well as the regulatory soluble synapse-specific protease Neurotrypsin. In this review we summarize the current state of the art regarding molecular structures and (agrin-dependent) canonical, as well as (agrin-independent) non-canonical, MuSK signaling mechanisms that underscore the formation of neuromuscular junctions, with the aim of providing a broad perspective to further stimulate molecular, cellular and tissue biology investigations on this fundamental intercellular contact.
Keywords: LRP4; MuSK; acetylcholine receptor; agrin; neuromuscular junctions; neurotrypsin; tyrosine kinase signaling.
Copyright © 2020 Guarino, Canciani and Forneris.
Figures
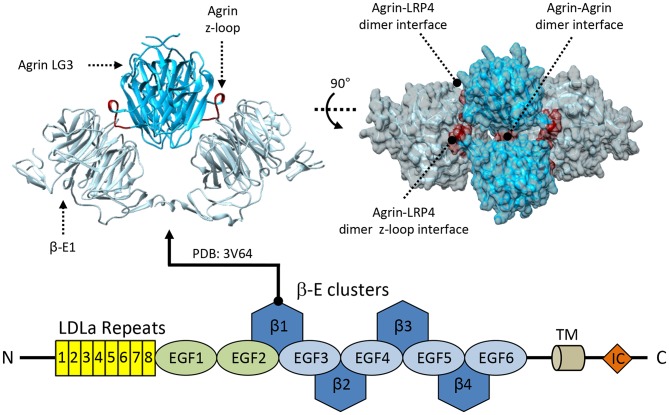
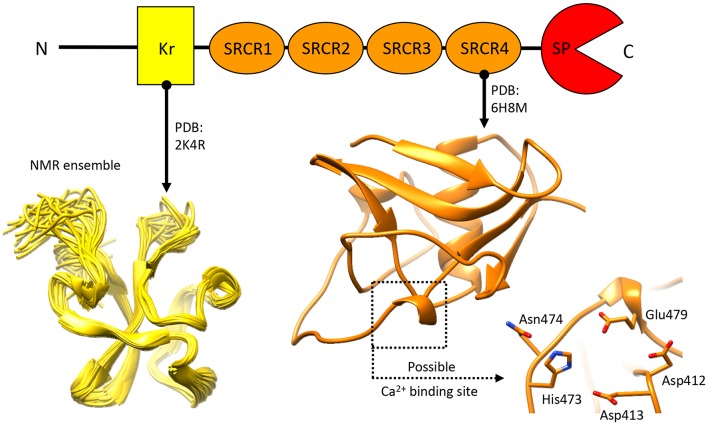
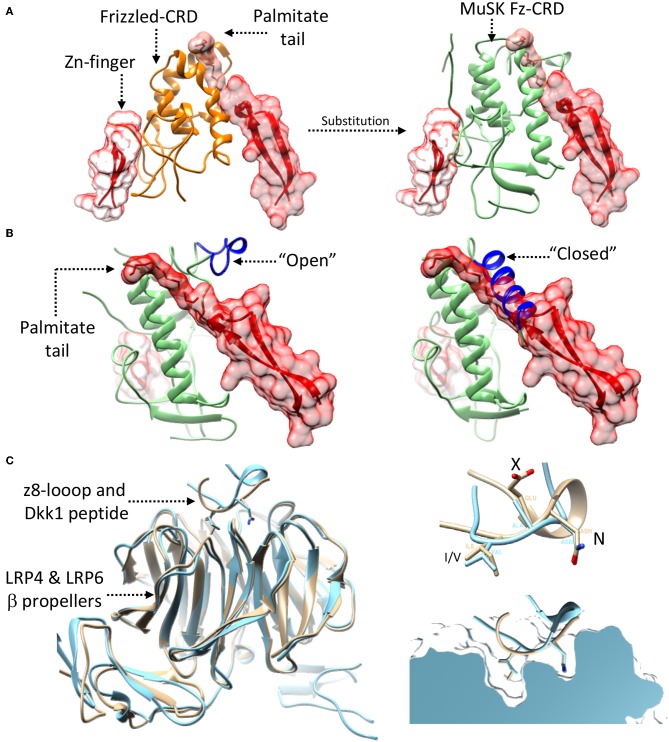
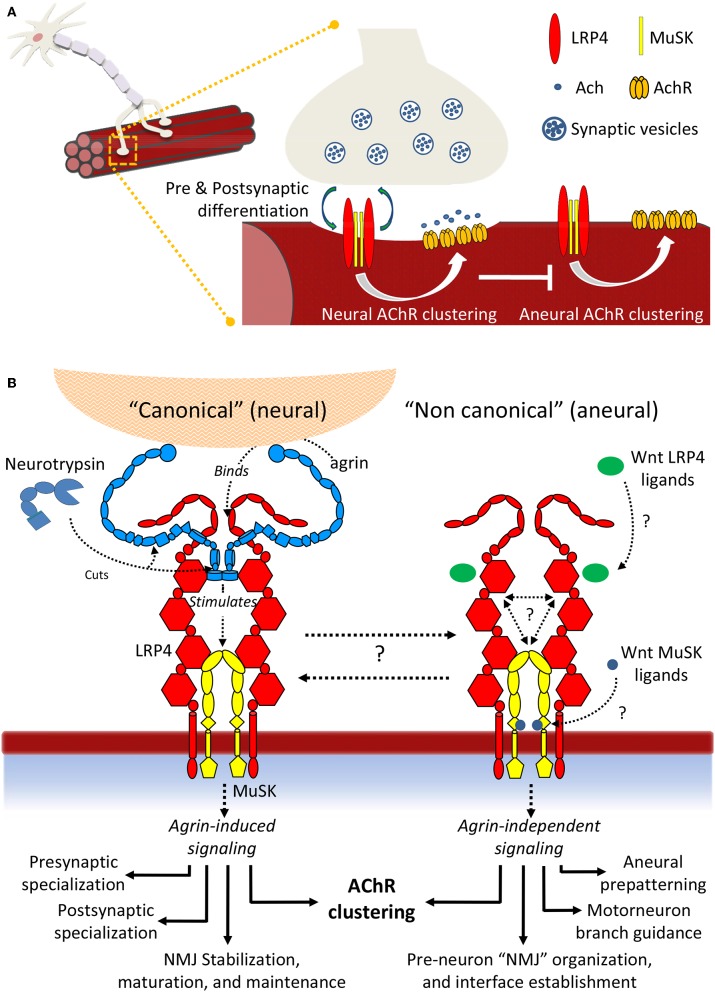
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous