

The noncanonical small heat shock protein HSP-17 from Caenorhabditis elegans is a selective protein aggregase
- PMID: 32001616
- PMCID: PMC7062175
- DOI: 10.1074/jbc.RA119.011185
The noncanonical small heat shock protein HSP-17 from Caenorhabditis elegans is a selective protein aggregase
Abstract
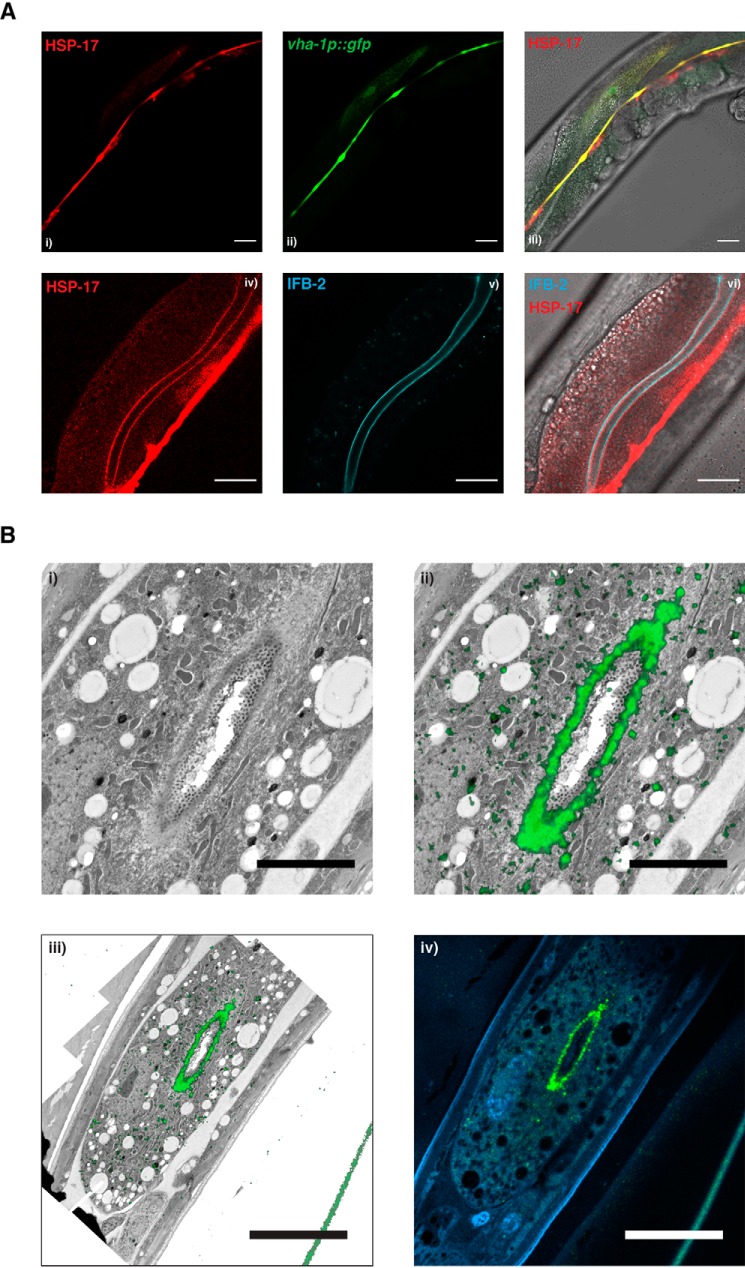
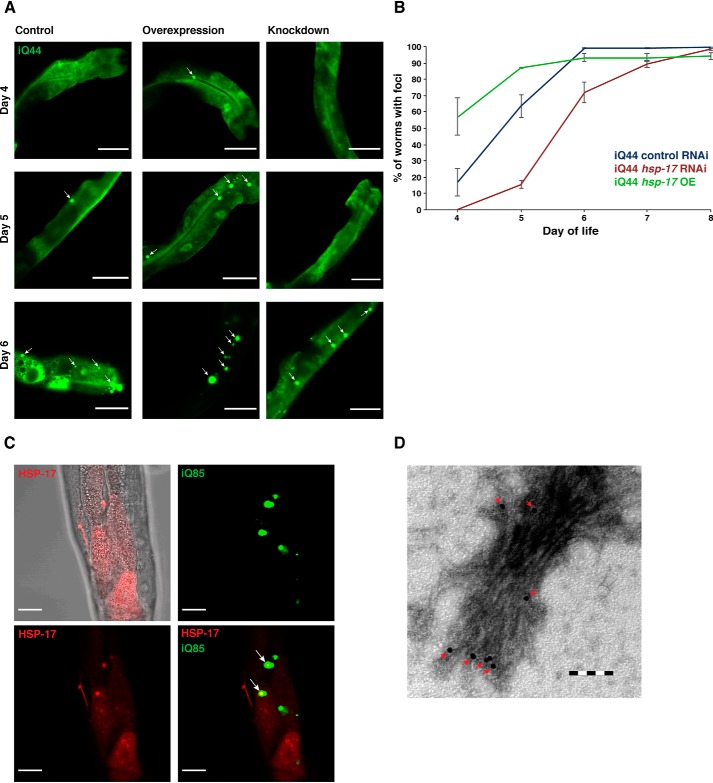
Small heat shock proteins (sHsps) are conserved, ubiquitous members of the proteostasis network. Canonically, they act as "holdases" and buffer unfolded or misfolded proteins against aggregation in an ATP-independent manner. Whereas bacteria and yeast each have only two sHsps in their genomes, this number is higher in metazoan genomes, suggesting a spatiotemporal and functional specialization in higher eukaryotes. Here, using recombinantly expressed and purified proteins, static light-scattering analysis, and disaggregation assays, we report that the noncanonical sHsp HSP-17 of Caenorhabditis elegans facilitates aggregation of model substrates, such as malate dehydrogenase (MDH), and inhibits disaggregation of luciferase in vitro Experiments with fluorescently tagged HSP-17 under the control of its endogenous promoter revealed that HSP-17 is expressed in the digestive and excretory organs, where its overexpression promotes the aggregation of polyQ proteins and of the endogenous kinase KIN-19. Systemic depletion of hsp-17 shortens C. elegans lifespan and severely reduces fecundity and survival upon prolonged heat stress. HSP-17 is an abundant protein exhibiting opposing chaperone activities on different substrates, indicating that it is a selective protein aggregase with physiological roles in development, digestion, and osmoregulation.
Keywords: Caenorhabditis elegans (C. elegans); chaperone; protein aggregates; protein aggregation; protein folding; proteostasis; selective protein aggregase; small heat shock protein (sHsp).
© 2020 Iburg et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
