

Core and Differentially Abundant Bacterial Taxa in the Rhizosphere of Field Grown Brassica napus Genotypes: Implications for Canola Breeding
- PMID: 32010086
- PMCID: PMC6974584
- DOI: 10.3389/fmicb.2019.03007
Core and Differentially Abundant Bacterial Taxa in the Rhizosphere of Field Grown Brassica napus Genotypes: Implications for Canola Breeding
Abstract
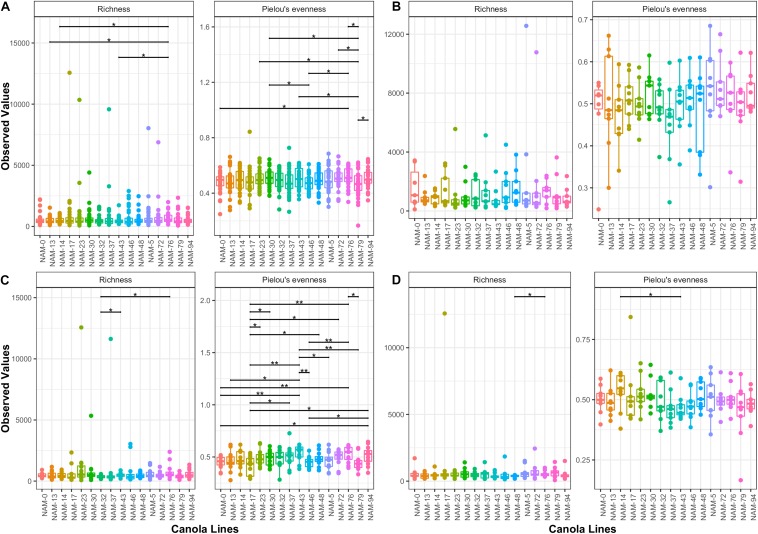
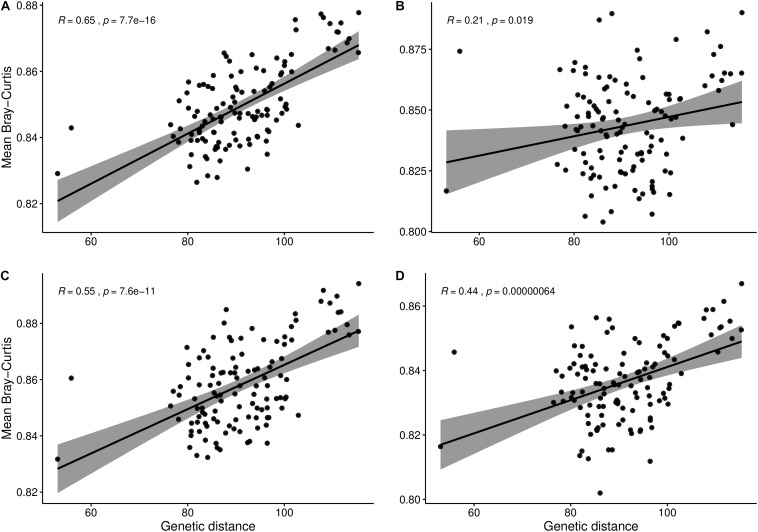
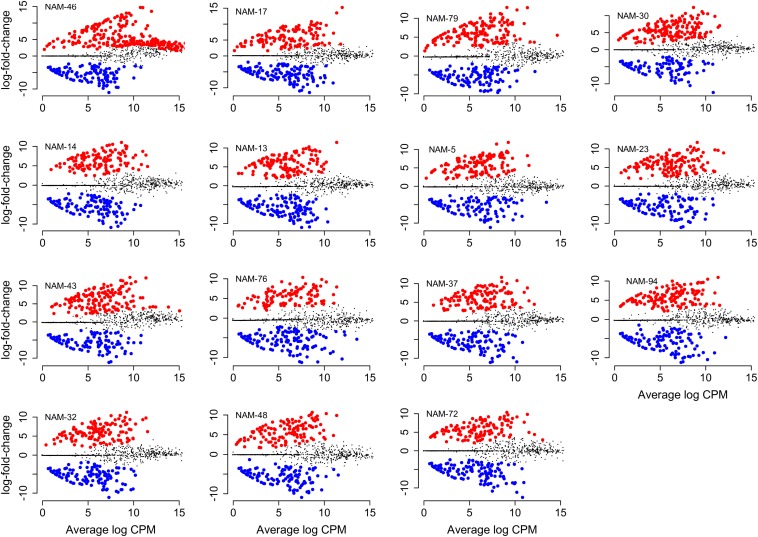
Modifying the rhizosphere microbiome through targeted plant breeding is key to harnessing positive plant-microbial interrelationships in cropping agroecosystems. Here, we examine the composition of rhizosphere bacterial communities of diverse Brassica napus genotypes to identify: (1) taxa that preferentially associate with genotypes, (2) core bacterial microbiota associated with B. napus, (3) heritable alpha diversity measures at flowering and whole growing season, and (4) correlation between microbial and plant genetic distance among canola genotypes at different growth stages. Our aim is to identify and describe signature microbiota with potential positive benefits that could be integrated in B. napus breeding and management strategies. Rhizosphere soils of 16 diverse genotypes sampled weekly over a 10-week period at single location as well as at three time points at two additional locations were analyzed using 16S rRNA gene amplicon sequencing. The B. napus rhizosphere microbiome was characterized by diverse bacterial communities with 32 named bacterial phyla. The most abundant phyla were Proteobacteria, Actinobacteria, and Acidobacteria. Overall microbial and plant genetic distances were highly correlated (R = 0.65). Alpha diversity heritability estimates were between 0.16 and 0.41 when evaluated across growth stage and between 0.24 and 0.59 at flowering. Compared with a reference B. napus genotype, a total of 81 genera were significantly more abundant and 71 were significantly less abundant in at least one B. napus genotype out of the total 558 bacterial genera. Most differentially abundant genera were Proteobacteria and Actinobacteria followed by Bacteroidetes and Firmicutes. Here, we also show that B. napus genotypes select an overall core bacterial microbiome with growth-stage-related patterns as to how taxa joined the core membership. In addition, we report that sets of B. napus core taxa were consistent across our three sites and 2 years. Both differential abundance and core analysis implicate numerous bacteria that have been reported to have beneficial effects on plant growth including disease suppression, antifungal properties, and plant growth promotion. Using a multi-site year, temporally intensive field sampling approach, we showed that small plant genetic differences cause predictable changes in canola microbiome and are potential target for direct and indirect selection within breeding programs.
Keywords: Brassica napus; breeding; canola; core microbiome; differential abundance; microbiome; plant–microbial interactions; rhizosphere.
Copyright © 2020 Taye, Helgason, Bell, Norris, Vail, Robinson, Parkin, Arcand, Mamet, Links, Dowhy, Siciliano and Lamb.
Figures
References
-
- Almario J., Kyselková M., Kopecký J., Ságová-Marečková M., Muller D., Grundmann G. L., et al. (2013). Assessment of the relationship between geologic origin of soil, rhizobacterial community composition and soil receptivity to tobacco black root rot in Savoie region (France). Plant Soil 371 397–408. 10.1007/s11104-013-1677-71 - DOI
-
- Alström S. (2001). Characteristics of bacteria from oilseed rape in relation to their biocontrol activity against Verticillium dahliae. J. Phytopathol. 149 57–64. 10.1046/j.1439-0434.2001.00585.x - DOI
-
- Anderson M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 26 32–46. 10.1111/j.1442-9993.2001.01070.pp.x - DOI
LinkOut - more resources
Full Text Sources