

Mutational processes of distinct POLE exonuclease domain mutants drive an enrichment of a specific TP53 mutation in colorectal cancer
- PMID: 32012149
- PMCID: PMC7018097
- DOI: 10.1371/journal.pgen.1008572
Mutational processes of distinct POLE exonuclease domain mutants drive an enrichment of a specific TP53 mutation in colorectal cancer
Abstract
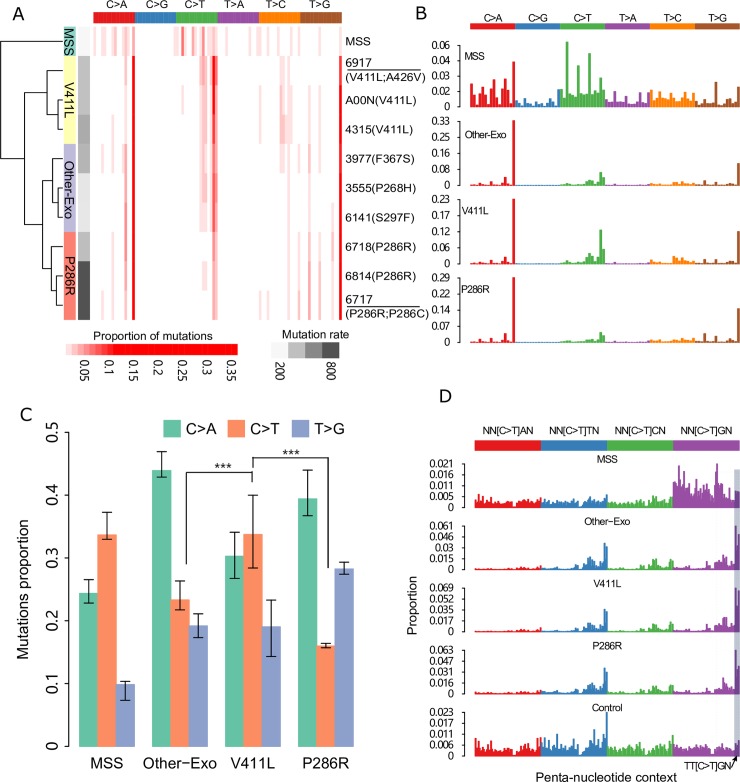
Cancer genomes with mutations in the exonuclease domain of Polymerase Epsilon (POLE) present with an extraordinarily high somatic mutation burden. In vitro studies have shown that distinct POLE mutants exhibit different polymerase activity. Yet, genome-wide mutation patterns and driver mutation formation arising from different POLE mutants remains unclear. Here, we curated somatic mutation calls from 7,345 colorectal cancer samples from published studies and publicly available databases. These include 44 POLE mutant samples including 9 with whole genome sequencing data available. The POLE mutant samples were categorized based on the specific POLE mutation present. Mutation spectrum, associations of somatic mutations with epigenomics features and co-occurrence with specific driver mutations were examined across different POLE mutants. We found that different POLE mutants exhibit distinct mutation spectrum with significantly higher relative frequency of C>T mutations in POLE V411L mutants. Our analysis showed that this increase frequency in C>T mutations is not dependent on DNA methylation and not associated with other genomic features and is thus specifically due to DNA sequence context alone. Notably, we found strong association of the TP53 R213* mutation specifically with POLE P286R mutants. This truncation mutation occurs within the TT[C>T]GA context. For C>T mutations, this sequence context is significantly more likely to be mutated in POLE P286R mutants compared with other POLE exonuclease domain mutants. This study refines our understanding of DNA polymerase fidelity and underscores genome-wide mutation spectrum and specific cancer driver mutation formation observed in POLE mutant cancers.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Jansen A.M., van Wezel T., van den Akker B.E., Ventayol Garcia M., Ruano D., Tops C.M., Wagner A., Letteboer T.G., Gomez-Garcia E.B., Devilee P. et al. (2016) Combined mismatch repair and POLE/POLD1 defects explain unresolved suspected Lynch syndrome cancers. Eur J Hum Genet, 24, 1089–1092. 10.1038/ejhg.2015.252 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
