

Aedes cadherin receptor that mediates Bacillus thuringiensis Cry11A toxicity is essential for mosquito development
- PMID: 32012156
- PMCID: PMC7018227
- DOI: 10.1371/journal.pntd.0007948
Aedes cadherin receptor that mediates Bacillus thuringiensis Cry11A toxicity is essential for mosquito development
Abstract
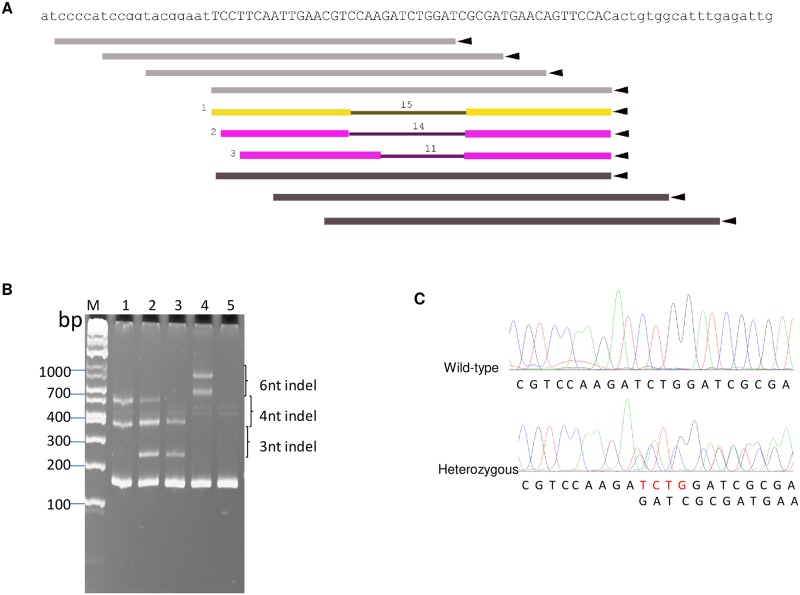
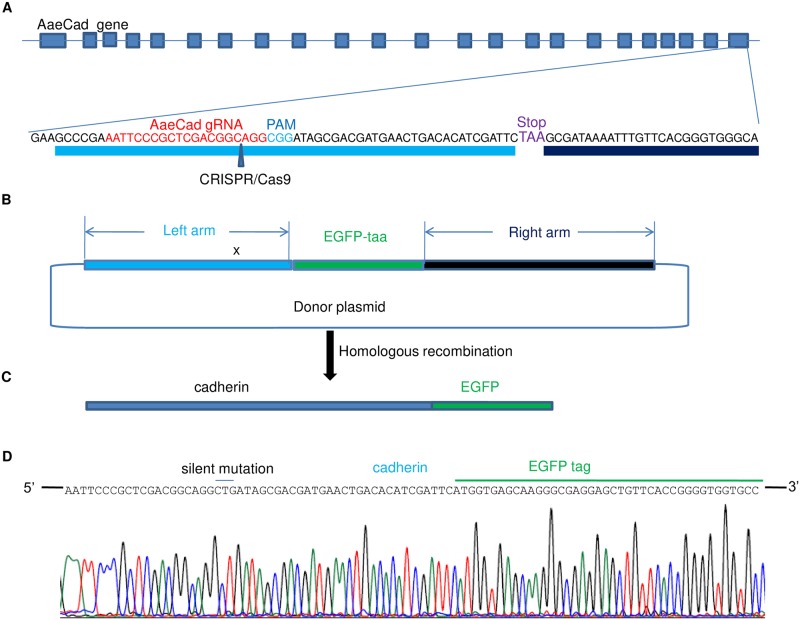
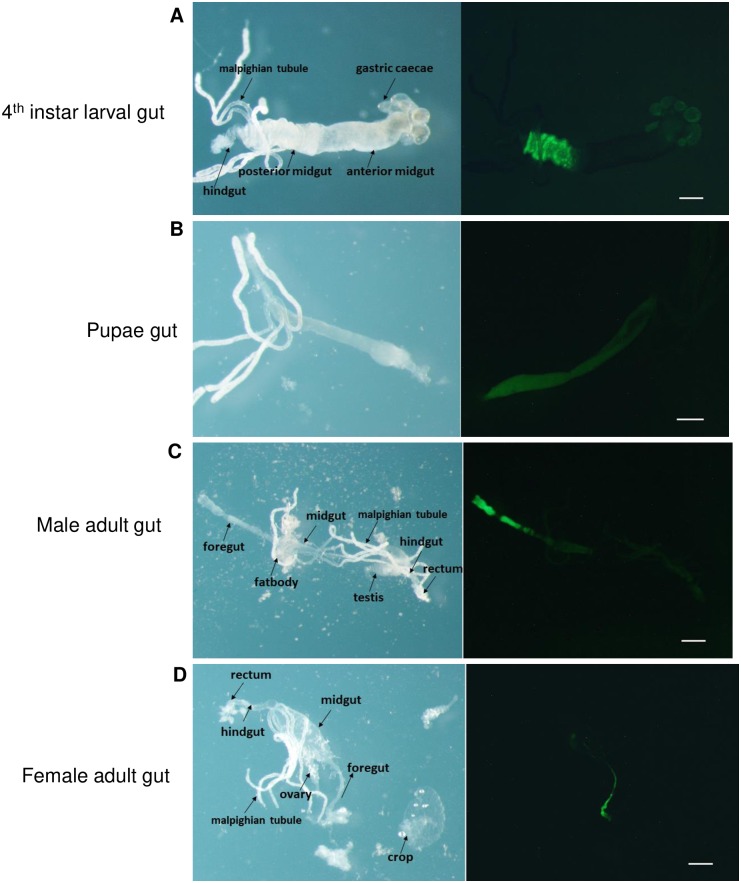
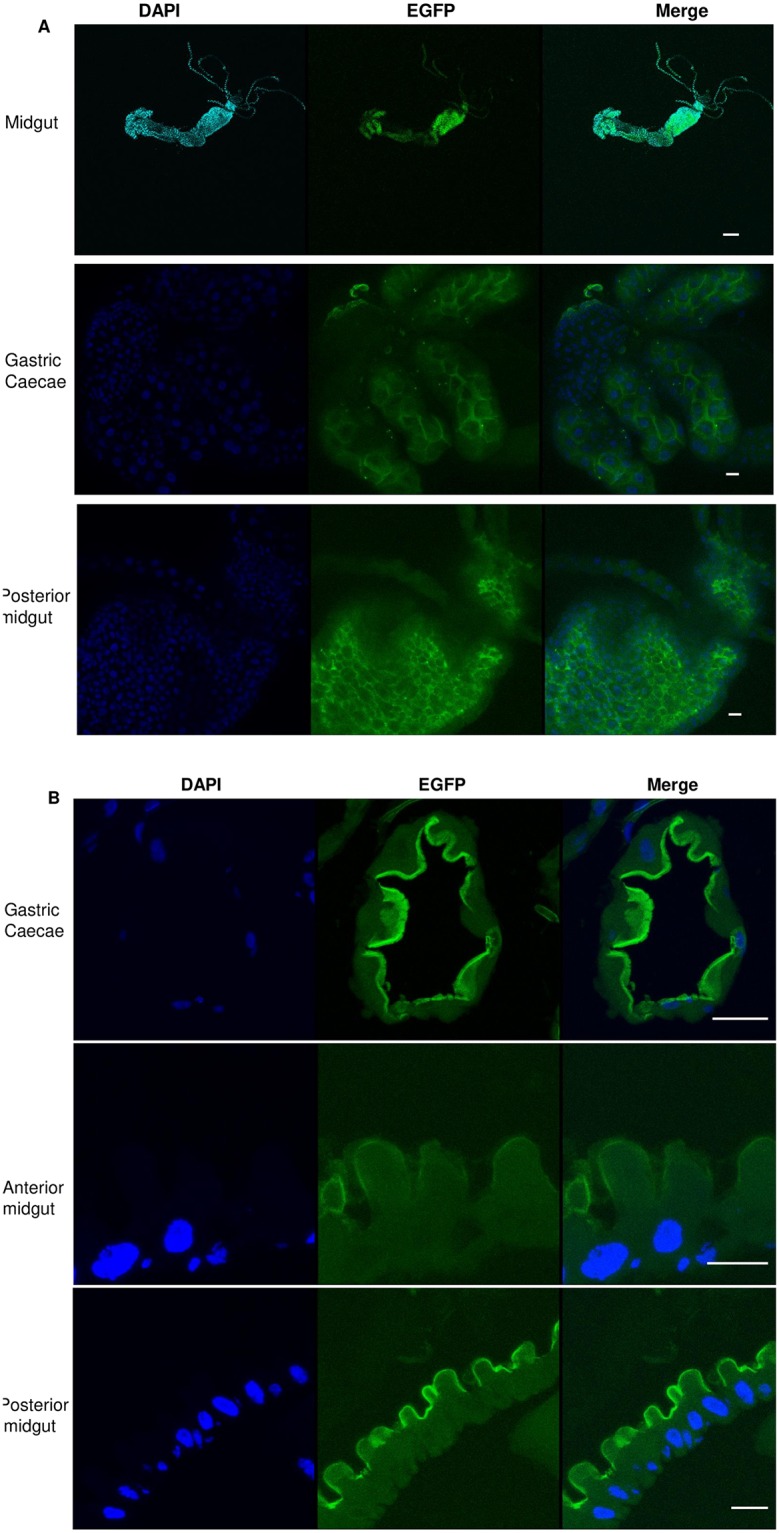
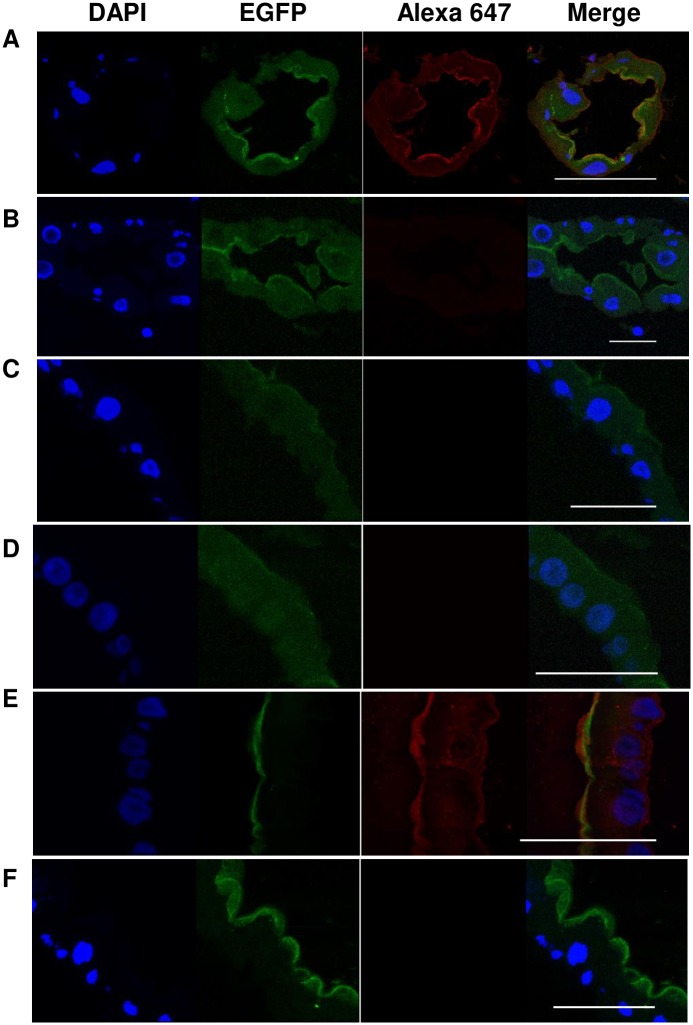
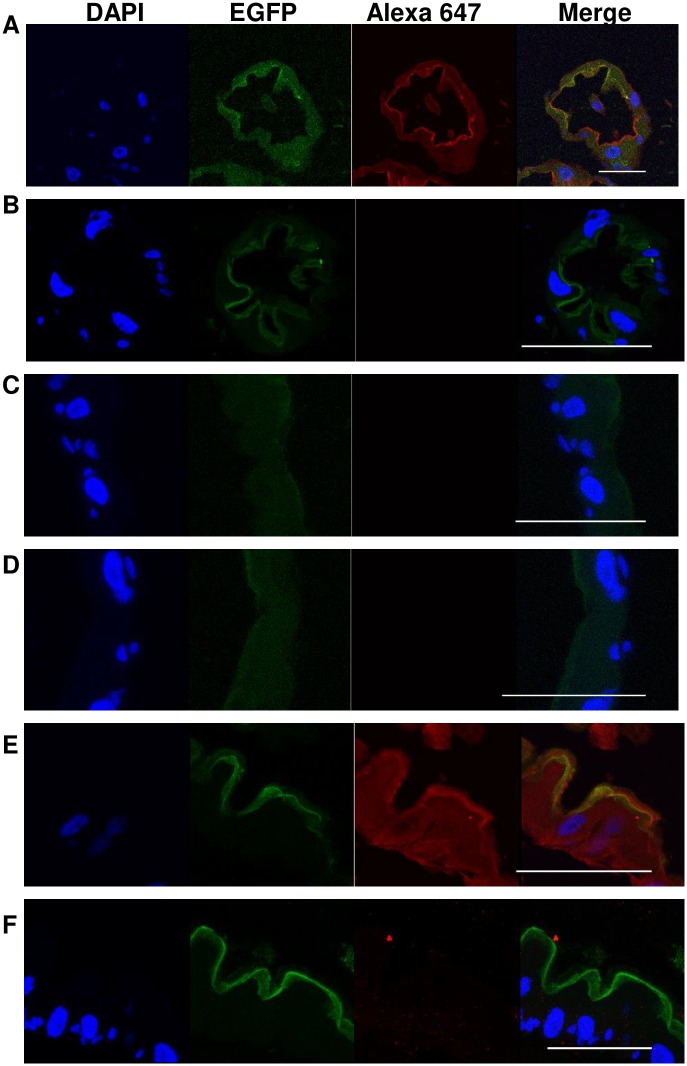
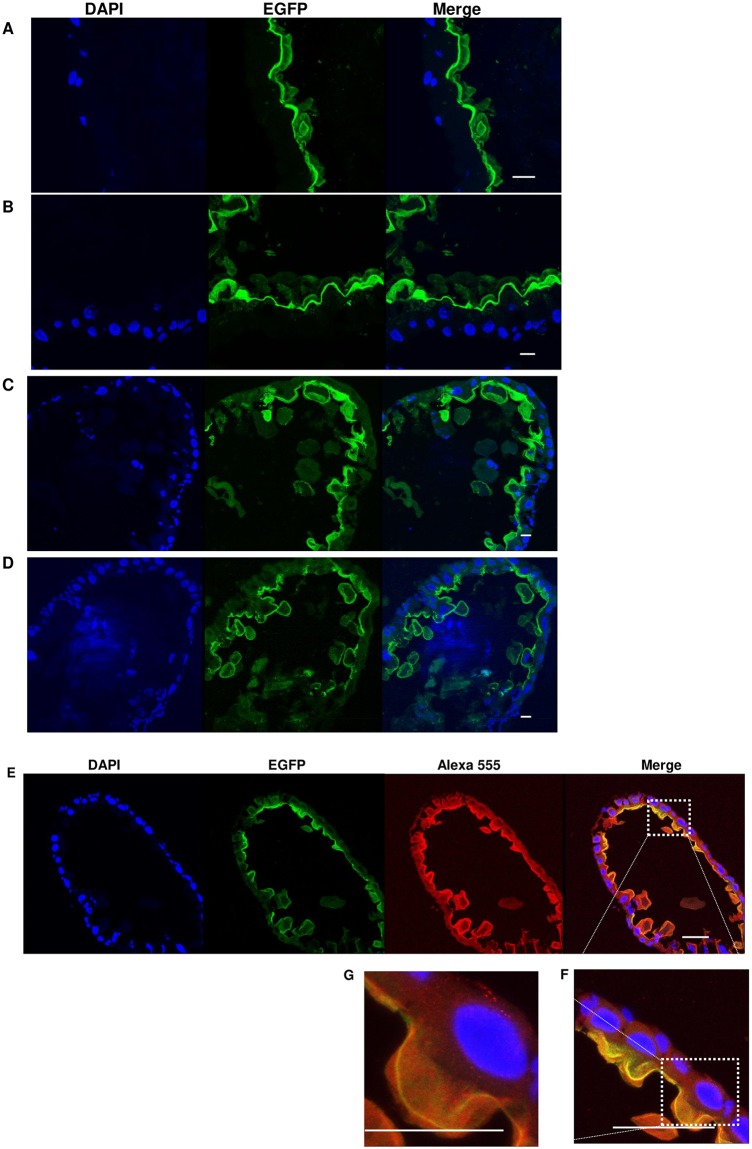
Aedes cadherin (AaeCad, AAEL024535) has been characterized as a receptor for Bacillus thuringiensis subsp. israelensis (Bti) Cry11A toxins. However, its role in development is still unknown. In this study, we modified the cadherin gene using ZFN and TALEN. Even though we obtained heterozygous deletions, no homozygous mutants were viable. Because ZFN and TALEN have lower off-targets than CRISPR/Cas9, we conclude the cadherin gene is essential for Aedes development. In contrast, in lepidopteran insects loss of a homologous cadherin does not appear to be lethal, since homozygous mutants are viable. To analyze the role of AaeCad in vivo, we tagged this protein with EGFP using CRISPR-Cas9-mediated homologous recombination and obtained a homozygous AaeCad-EGFP line. Addition of Aedes Rad51 mRNA enhanced the rate of recombination. We then examined AaeCad protein expression in most tissues and protein dynamics during mosquito development. We observe that AaeCad is expressed in larval and adult midgut-specific manner and its expression pattern changed during the mosquito development. Confocal images showed AaeCad has high expression in larval caecae and posterior midgut, and also in adult midgut. Expression of AaeCad is observed primarily in the apical membranes of epithelial cells, and not in cell-cell junctions. The expression pattern observed suggests AaeCad does not appear to play a role in these junctions. However, we cannot exclude its role beyond cell-cell adhesion in the midgut. We also observed that Cry11A bound to the apical side of larval gastric caecae and posterior midgut cells exactly where AaeCad-EGFP was expressed. Their co-localization suggests that AaeCad is indeed a receptor for the Cry11A toxin. Using this mosquito line we also observed that low doses of Cry11A toxin caused the cells to slough off membranes, which likely represents a defense mechanism, to limit cell damage from Cry11A toxin pores formed in the cell membrane.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
