

ALT: A Multi-Faceted Phenomenon
- PMID: 32012790
- PMCID: PMC7073516
- DOI: 10.3390/genes11020133
ALT: A Multi-Faceted Phenomenon
Abstract
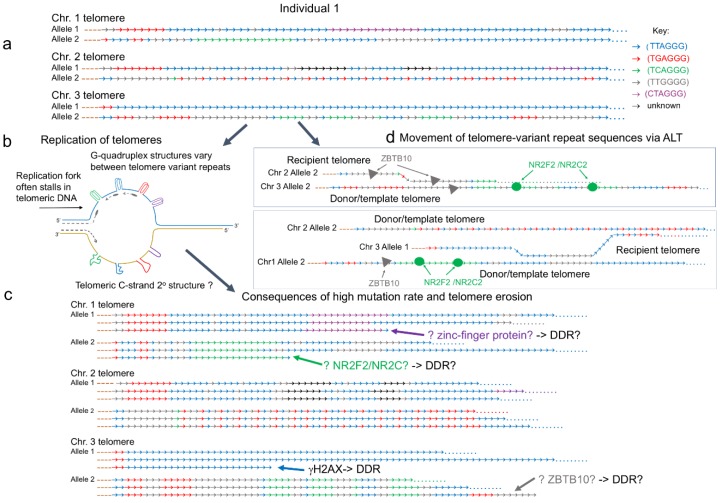
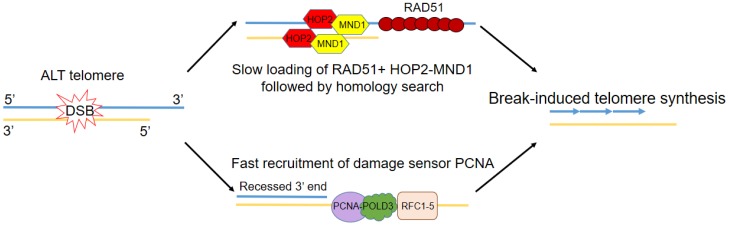
One of the hallmarks of cancer cells is their indefinite replicative potential, made possible by the activation of a telomere maintenance mechanism (TMM). The majority of cancers reactivate the reverse transcriptase, telomerase, to maintain their telomere length but a minority (10% to 15%) utilize an alternative lengthening of telomeres (ALT) pathway. Here, we review the phenotypes and molecular markers specific to ALT, and investigate the significance of telomere mutations and sequence variation in ALT cell lines. We also look at the recent advancements in understanding the different mechanisms behind ALT telomere elongation and finally, the progress made in identifying potential ALT-targeted therapies, including those already in use for the treatment of both hematological and solid tumors.
Keywords: alternative lengthening of telomeres; break-induced telomere synthesis; cancer; telomere-variant-repeat; therapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
