

Reciprocal Regulatory Interaction between TRPV1 and Kinin B1 Receptor in a Rat Neuropathic Pain Model
- PMID: 32012798
- PMCID: PMC7037982
- DOI: 10.3390/ijms21030821
Reciprocal Regulatory Interaction between TRPV1 and Kinin B1 Receptor in a Rat Neuropathic Pain Model
Abstract
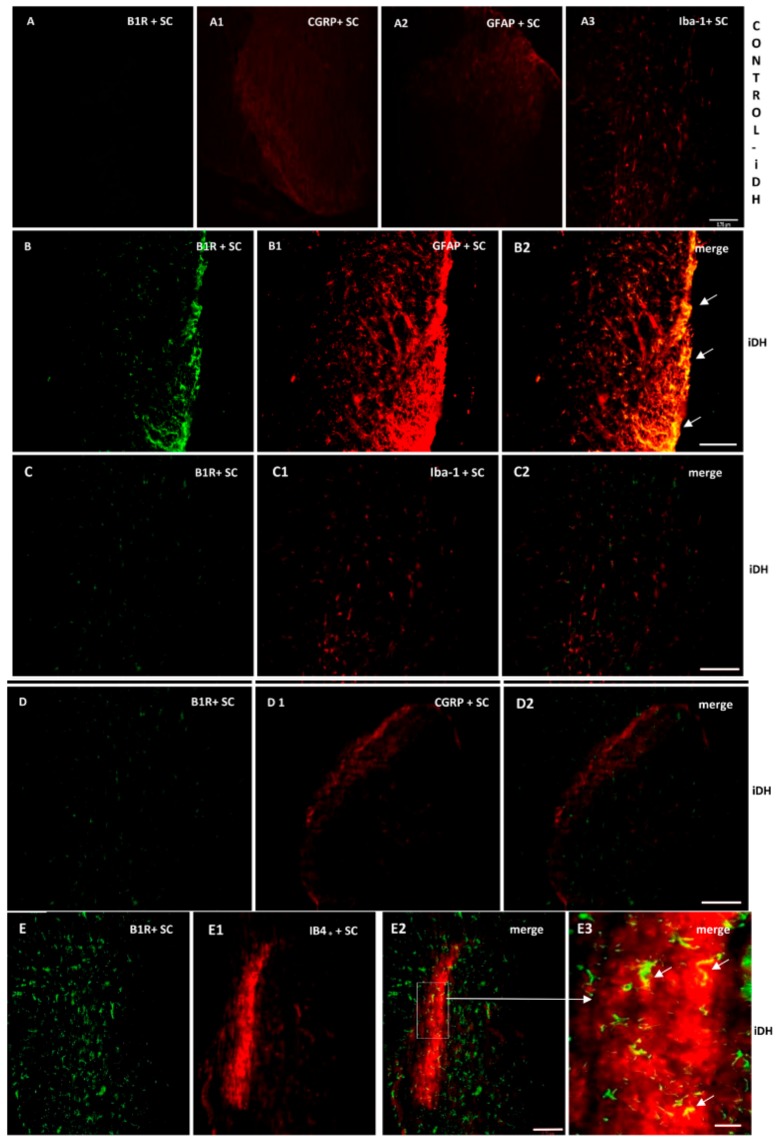
Kinins are mediators of pain and inflammation and evidence suggests that the inducible kinin B1 receptor (B1R) is involved in neuropathic pain (NP). This study investigates whether B1R and TRPV1 are colocalized on nociceptors and/or astrocytes to enable regulatory interaction either directly or through the cytokine pathway (IL-1β, TNF-α) in NP. Sprague Dawley rats were subjected to unilateral partial sciatic nerve ligation (PSNL) and treated from 14 to 21 days post-PSNL with antagonists of B1R (SSR240612, 10 mg·kg-1, i.p.) or TRPV1 (SB366791, 1 mg·kg-1, i.p.). The impact of these treatments was assessed on nociceptive behavior and mRNA expression of B1R, TRPV1, TNF-α, and IL-1β. Localization on primary sensory fibers, astrocytes, and microglia was determined by immunofluorescence in the lumbar spinal cord and dorsal root ganglion (DRG). Both antagonists suppressed PSNL-induced thermal hyperalgesia, but only SB366791 blunted mechanical and cold allodynia. SSR240612 reversed PSNL-induced enhanced protein and mRNA expression of B1R and TRPV1 mRNA levels in spinal cord while SB366791 further increased B1R mRNA/protein expression. B1R and TRPV1 were found in non-peptide sensory fibers and astrocytes, and colocalized in the spinal dorsal horn and DRG, notably with IL-1β on astrocytes. IL-1β mRNA further increased under B1R or TRPV1 antagonism. Data suggest that B1R and TRPV1 contribute to thermal hyperalgesia and play a distinctive role in allodynia associated with NP. Close interaction and reciprocal regulatory mechanism are suggested between B1R and TRPV1 on astrocytes and nociceptors in NP.
Keywords: B1 receptor; TRPV1; allodynia; astrocytes; bradykinin; cytokines; dorsal root ganglion; hyperalgesia; sensory fibers; spinal cord.
Conflict of interest statement
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures








References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
