

A thermophilic phage uses a small terminase protein with a fixed helix-turn-helix geometry
- PMID: 32014998
- PMCID: PMC7086035
- DOI: 10.1074/jbc.RA119.012224
A thermophilic phage uses a small terminase protein with a fixed helix-turn-helix geometry
Abstract
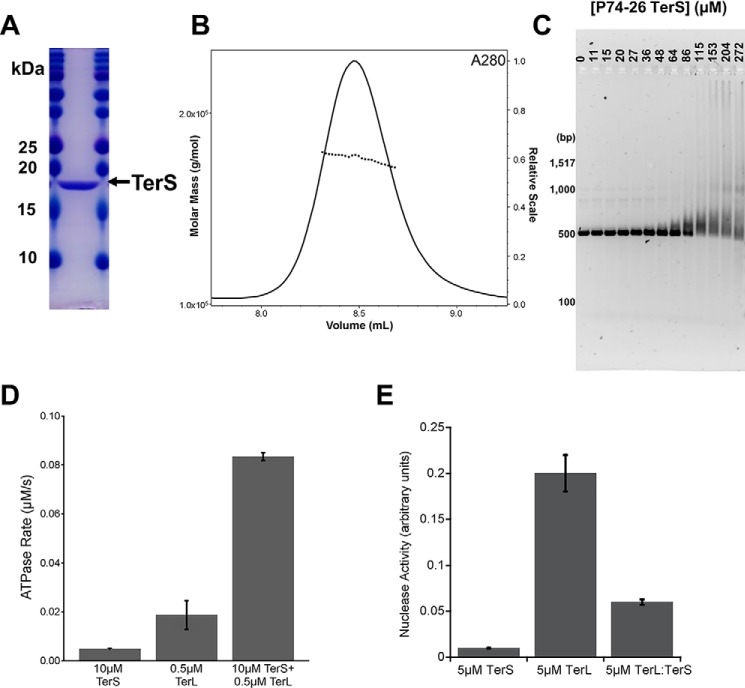
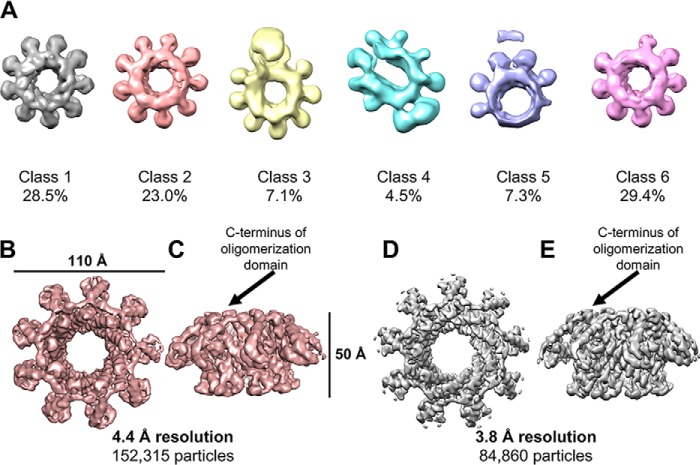
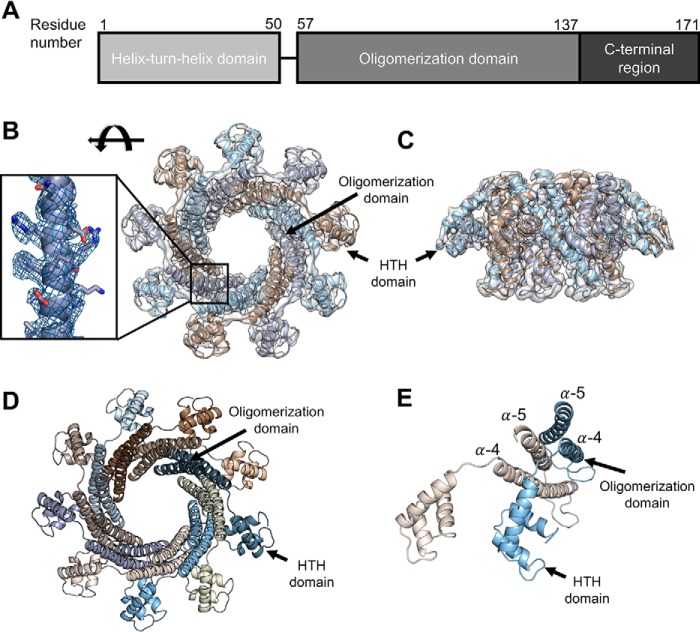
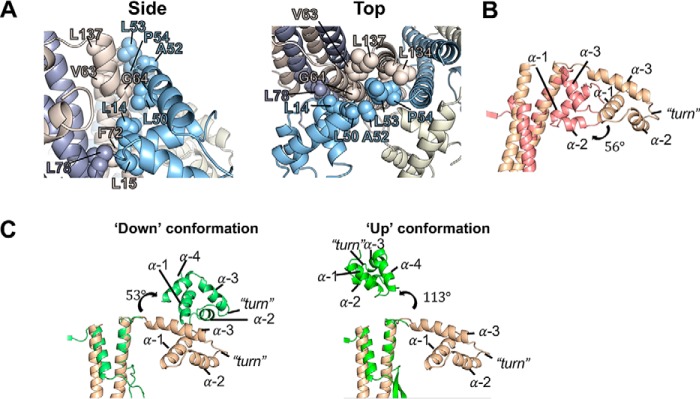
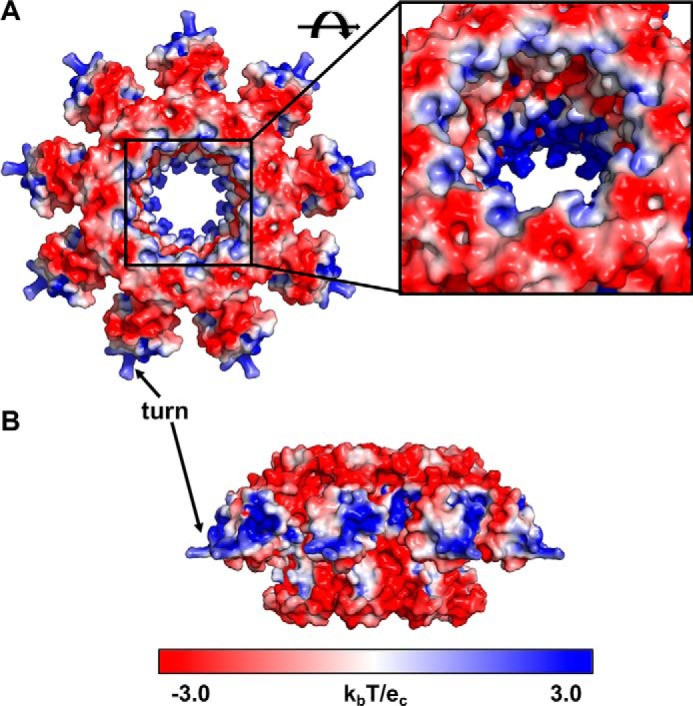
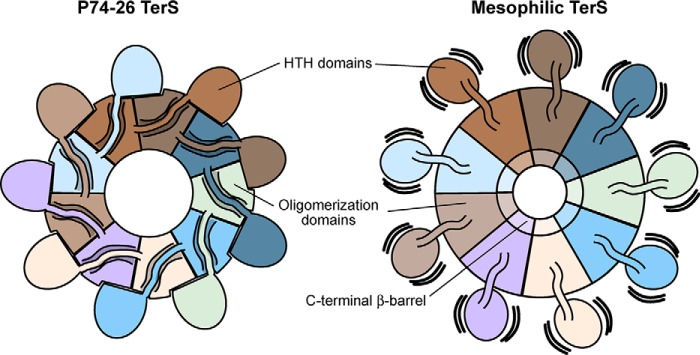
Tailed bacteriophages use a DNA-packaging motor to encapsulate their genome during viral particle assembly. The small terminase (TerS) component of this DNA-packaging machinery acts as a molecular matchmaker that recognizes both the viral genome and the main motor component, the large terminase (TerL). However, how TerS binds DNA and the TerL protein remains unclear. Here we identified gp83 of the thermophilic bacteriophage P74-26 as the TerS protein. We found that TerSP76-26 oligomerizes into a nonamer that binds DNA, stimulates TerL ATPase activity, and inhibits TerL nuclease activity. A cryo-EM structure of TerSP76-26 revealed that it forms a ring with a wide central pore and radially arrayed helix-turn-helix domains. The structure further showed that these helix-turn-helix domains, which are thought to bind DNA by wrapping the double helix around the ring, are rigidly held in an orientation distinct from that seen in other TerS proteins. This rigid arrangement of the putative DNA-binding domain imposed strong constraints on how TerSP76-26 can bind DNA. Finally, the TerSP76-26 structure lacked the conserved C-terminal β-barrel domain used by other TerS proteins for binding TerL. This suggests that a well-ordered C-terminal β-barrel domain is not required for TerSP76-26 to carry out its matchmaking function. Our work highlights a thermophilic system for studying the role of small terminase proteins in viral maturation and presents the structure of TerSP76-26, revealing key differences between this thermophilic phage and its mesophilic counterparts.
Keywords: DNA packaging; DNA recognition; DNA-binding protein; bacteriophage; cryo-EM; helix–turn–helix domain; molecular motor; small terminase; thermophile; viral motor.
© 2020 Hayes et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Feiss M., and Rao V. B. (2012) The Bacteriophage DNA Packaging Machine. Adv. Exp. Med. Biol. 726, 489–509 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
