

Interleukin-8 Dedifferentiates Primary Human Luminal Cells to Multipotent Stem Cells
- PMID: 32015100
- PMCID: PMC7156216
- DOI: 10.1128/MCB.00508-19
Interleukin-8 Dedifferentiates Primary Human Luminal Cells to Multipotent Stem Cells
Abstract
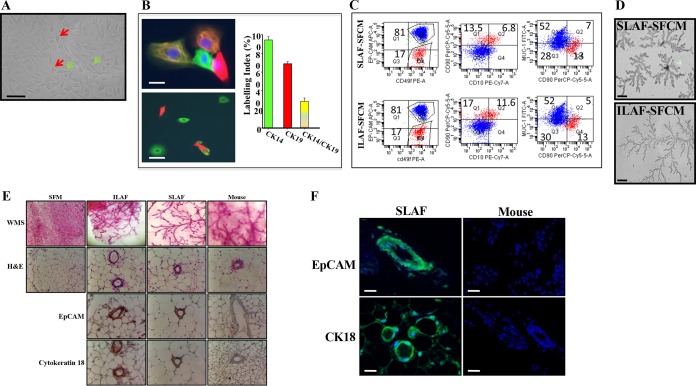
During aging, cellular plasticity and senescence play important roles in tissue regeneration and the pathogenesis of different diseases, including cancer. We have recently shown that senescent breast luminal cells can activate their adjacent stromal fibroblasts. In the present report, we present clear evidence that these senescence-related active fibroblasts can dedifferentiate proliferating primary human luminal cells to multipotent stem cells in an interleukin-8 (IL-8)-dependent manner. This was confirmed using recombinant IL-8, while the truncated protein was not active. This IL-8-related dedifferentiation of luminal cells was mediated through the STAT3-dependent downregulation of p16INK4A and the microRNA miR-141. Importantly, these in vitro-generated mammary stem cells exhibited high molecular and cellular similarities to human mammary stem cells. They have also shown a long-term mammary gland-reconstituting ability and the capacity to produce milk postdelivery. Thereby, these IL-8-generated mammary stem cells could be of great value for autologous cell therapy procedures and also for biomedical research as well as drug development.
Keywords: IL-8; STAT3; cell differentiation; stem cells.
Copyright © 2020 American Society for Microbiology.
Figures










Similar articles
-
Senescent Breast Luminal Cells Promote Carcinogenesis through Interleukin-8-Dependent Activation of Stromal Fibroblasts.Mol Cell Biol. 2019 Jan 3;39(2):e00359-18. doi: 10.1128/MCB.00359-18. Print 2019 Jan 15. Mol Cell Biol. 2019. PMID: 30397077 Free PMC article.
-
miR-146b-5p mediates p16-dependent repression of IL-6 and suppresses paracrine procarcinogenic effects of breast stromal fibroblasts.Oncotarget. 2015 Oct 6;6(30):30006-16. doi: 10.18632/oncotarget.4933. Oncotarget. 2015. PMID: 26338965 Free PMC article.
-
The cytokine IL-6 reactivates breast stromal fibroblasts through transcription factor STAT3-dependent up-regulation of the RNA-binding protein AUF1.J Biol Chem. 2014 Nov 7;289(45):30962-76. doi: 10.1074/jbc.M114.594044. Epub 2014 Sep 17. J Biol Chem. 2014. PMID: 25231991 Free PMC article.
-
Concise review: MicroRNA expression in multipotent mesenchymal stromal cells.Stem Cells. 2008 Feb;26(2):356-63. doi: 10.1634/stemcells.2007-0625. Epub 2007 Nov 8. Stem Cells. 2008. PMID: 17991914 Free PMC article. Review.
-
Molecular changes accompanying senescence and immortalization of cultured human mammary epithelial cells.Int J Biochem Cell Biol. 2002 Nov;34(11):1382-94. doi: 10.1016/s1357-2725(02)00047-x. Int J Biochem Cell Biol. 2002. PMID: 12200033 Review.
Cited by
-
STAT3: Key targets of growth-promoting receptor positive breast cancer.Cancer Cell Int. 2024 Oct 28;24(1):356. doi: 10.1186/s12935-024-03541-9. Cancer Cell Int. 2024. PMID: 39468521 Free PMC article. Review.
-
A cell transcriptomic profile provides insights into adipocytes of porcine mammary gland across development.J Anim Sci Biotechnol. 2023 Oct 8;14(1):126. doi: 10.1186/s40104-023-00926-0. J Anim Sci Biotechnol. 2023. PMID: 37805503 Free PMC article.
-
Age-related and postmenopausal breast cancer progression and treatment management: The significance of pro-inflammatory cytokines and CXC chemokines.Genes Dis. 2025 Mar 19;12(5):101606. doi: 10.1016/j.gendis.2025.101606. eCollection 2025 Sep. Genes Dis. 2025. PMID: 40584290 Free PMC article. Review.
References
-
- Colacino JA, Azizi E, Brooks MD, Harouaka R, Fouladdel S, McDermott SP, Lee M, Hill D, Madden J, Boerner J, Cote ML, Sartor MA, Rozek LS, Wicha MS. 2018. Heterogeneity of human breast stem and progenitor cells as revealed by transcriptional profiling. Stem Cell Rep 10:1596–1609. doi:10.1016/j.stemcr.2018.03.001. - DOI - PMC - PubMed
-
- Milanovic M, Fan DNY, Belenki D, Dabritz JHM, Zhao Z, Yu Y, Dorr JR, Dimitrova L, Lenze D, Monteiro Barbosa IA, Mendoza-Parra MA, Kanashova T, Metzner M, Pardon K, Reimann M, Trumpp A, Dorken B, Zuber J, Gronemeyer H, Hummel M, Dittmar G, Lee S, Schmitt CA. 2018. Senescence-associated reprogramming promotes cancer stemness. Nature 553:96–100. doi:10.1038/nature25167. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous