

Synaptotagmin 1 oligomers clamp and regulate different modes of neurotransmitter release
- PMID: 32015138
- PMCID: PMC7035618
- DOI: 10.1073/pnas.1920403117
Synaptotagmin 1 oligomers clamp and regulate different modes of neurotransmitter release
Abstract
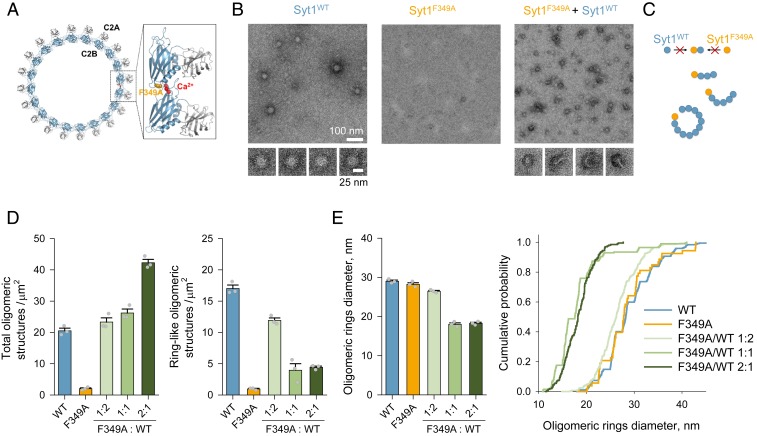
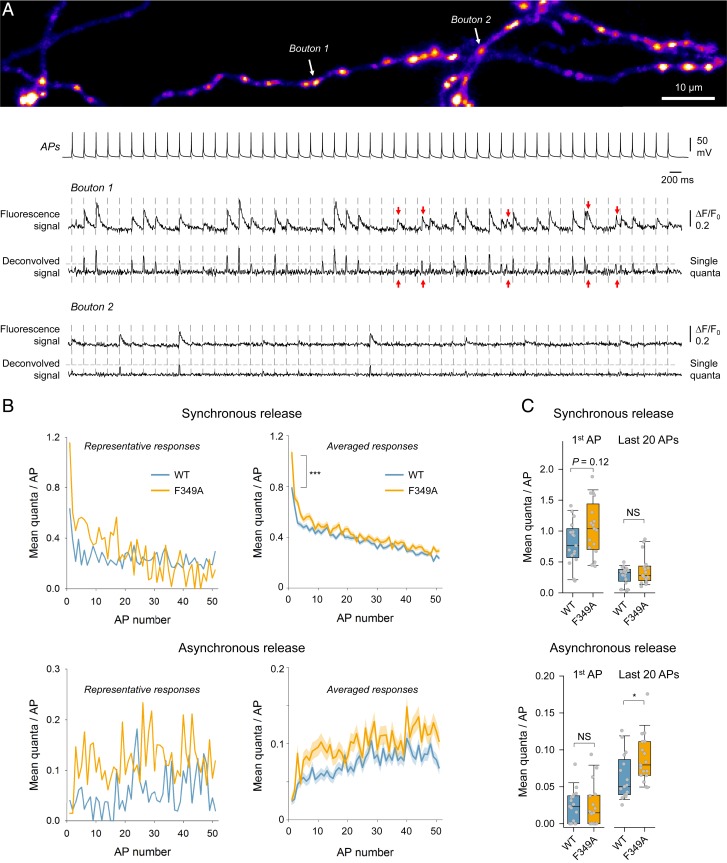
Synaptotagmin 1 (Syt1) synchronizes neurotransmitter release to action potentials (APs) acting as the fast Ca2+ release sensor and as the inhibitor (clamp) of spontaneous and delayed asynchronous release. While the Syt1 Ca2+ activation mechanism has been well-characterized, how Syt1 clamps transmitter release remains enigmatic. Here we show that C2B domain-dependent oligomerization provides the molecular basis for the Syt1 clamping function. This follows from the investigation of a designed mutation (F349A), which selectively destabilizes Syt1 oligomerization. Using a combination of fluorescence imaging and electrophysiology in neocortical synapses, we show that Syt1F349A is more efficient than wild-type Syt1 (Syt1WT) in triggering synchronous transmitter release but fails to clamp spontaneous and synaptotagmin 7 (Syt7)-mediated asynchronous release components both in rescue (Syt1-/- knockout background) and dominant-interference (Syt1+/+ background) conditions. Thus, we conclude that Ca2+-sensitive Syt1 oligomers, acting as an exocytosis clamp, are critical for maintaining the balance among the different modes of neurotransmitter release.
Keywords: C2B domain; fusion clamp; synaptic transmission; synaptotagmin.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures


References
-
- Schneggenburger R., Rosenmund C., Molecular mechanisms governing Ca(2+) regulation of evoked and spontaneous release. Nat. Neurosci. 18, 935–941 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
