

An optimized chemical-genetic method for cell-specific metabolic labeling of RNA
- PMID: 32015544
- PMCID: PMC8518020
- DOI: 10.1038/s41592-019-0726-y
An optimized chemical-genetic method for cell-specific metabolic labeling of RNA
Abstract
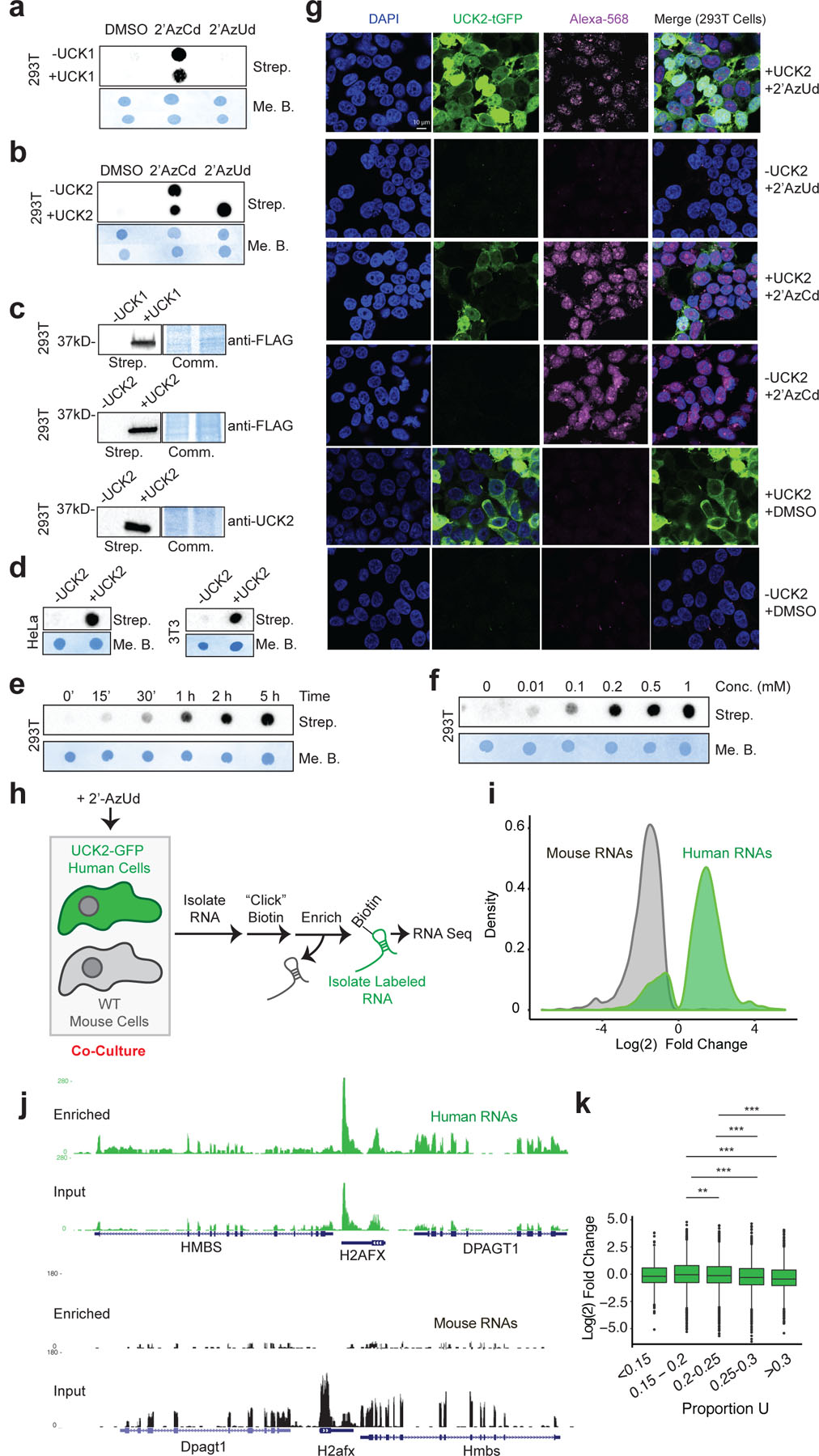
Tissues and organs are composed of diverse cell types, which poses a major challenge for cell-type-specific profiling of gene expression. Current metabolic labeling methods rely on exogenous pyrimidine analogs that are only incorporated into RNA in cells expressing an exogenous enzyme. This approach assumes that off-target cells cannot incorporate these analogs. We disprove this assumption and identify and characterize the enzymatic pathways responsible for high background incorporation. We demonstrate that mammalian cells can incorporate uracil analogs and characterize the enzymatic pathways responsible for high background incorporation. To overcome these limitations, we developed a new small molecule-enzyme pair consisting of uridine/cytidine kinase 2 and 2'-azidouridine. We demonstrate that 2'-azidouridine is only incorporated in cells expressing uridine/cytidine kinase 2 and characterize selectivity mechanisms using molecular dynamics and X-ray crystallography. Furthermore, this pair can be used to purify and track RNA from specific cellular populations, making it ideal for high-resolution cell-specific RNA labeling. Overall, these results reveal new aspects of mammalian salvage pathways and serve as a new benchmark for designing, characterizing and evaluating methodologies for cell-specific labeling of biomolecules.
Figures




References
References (Main Text)
-
- Jung J & Jung H. Methods to analyze cell type-specific gene expression profiles from heterogeneous cell populations. Anim Cells Syst 20, 113–117, doi: 10.1080/19768354.2016.1191544 (2016). - DOI
References (Methods)
-
- Agarwal KC, Miech RP & Parks RE in Methods Enzymol. Vol. 51 483–490 (Academic Press, 1978). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T34 GM069337/GM/NIGMS NIH HHS/United States
- 1DP2GM119164/U.S. Department of Health & Human Services | National Institutes of Health (NIH)/International
- R01 GM108889/GM/NIGMS NIH HHS/United States
- DP2 GM119164/GM/NIGMS NIH HHS/United States
- S10 RR025496/RR/NCRR NIH HHS/United States
- R21 MH113062/MH/NIMH NIH HHS/United States
- S10 OD010794/OD/NIH HHS/United States
- P30 CA062203/CA/NCI NIH HHS/United States
- 5R21MH113062/U.S. Department of Health & Human Services | National Institutes of Health (NIH)/International
- R25 GM055246/GM/NIGMS NIH HHS/United States
- S10 OD021718/OD/NIH HHS/United States
- 1R01GM108889-01/U.S. Department of Health & Human Services | National Institutes of Health (NIH)/International
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
