

Atomic structures of closed and open influenza B M2 proton channel reveal the conduction mechanism
- PMID: 32015551
- PMCID: PMC7641042
- DOI: 10.1038/s41594-019-0371-2
Atomic structures of closed and open influenza B M2 proton channel reveal the conduction mechanism
Abstract
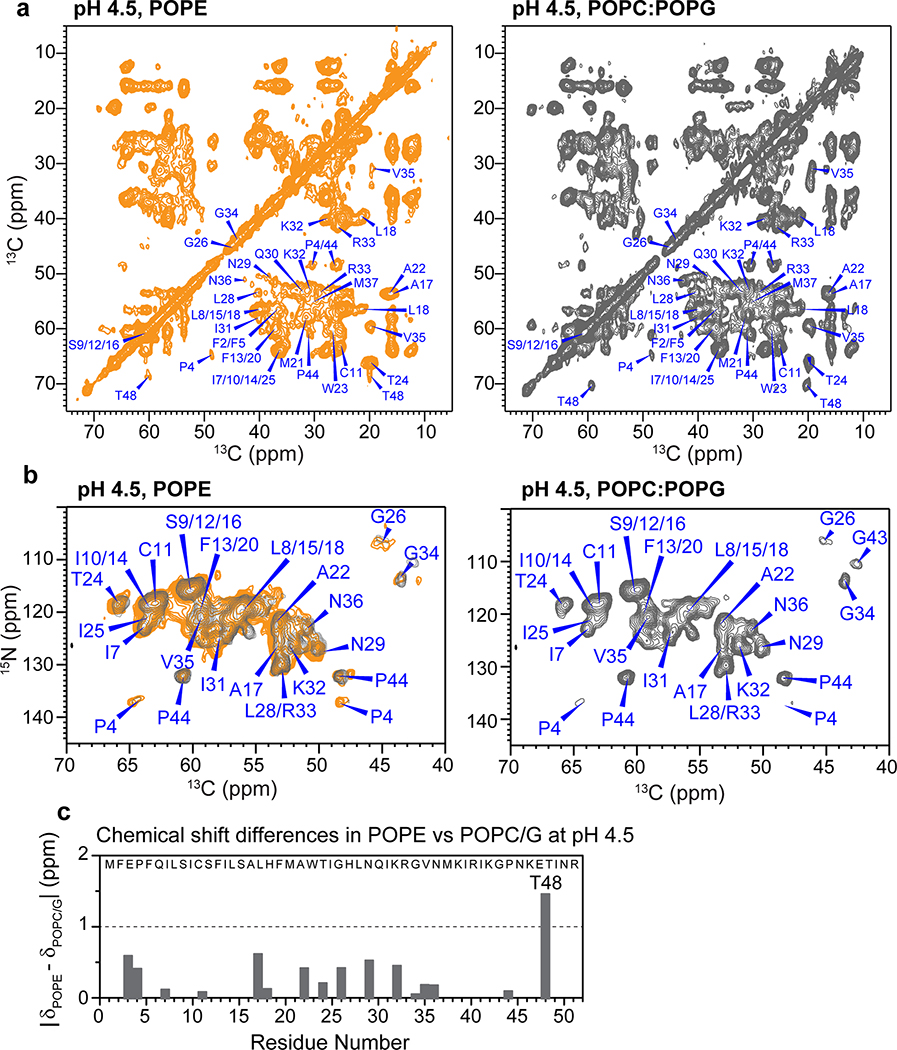
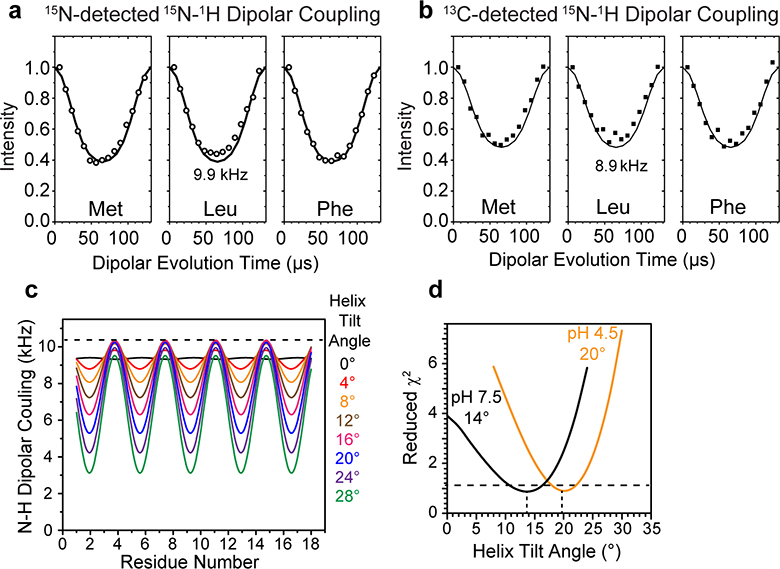
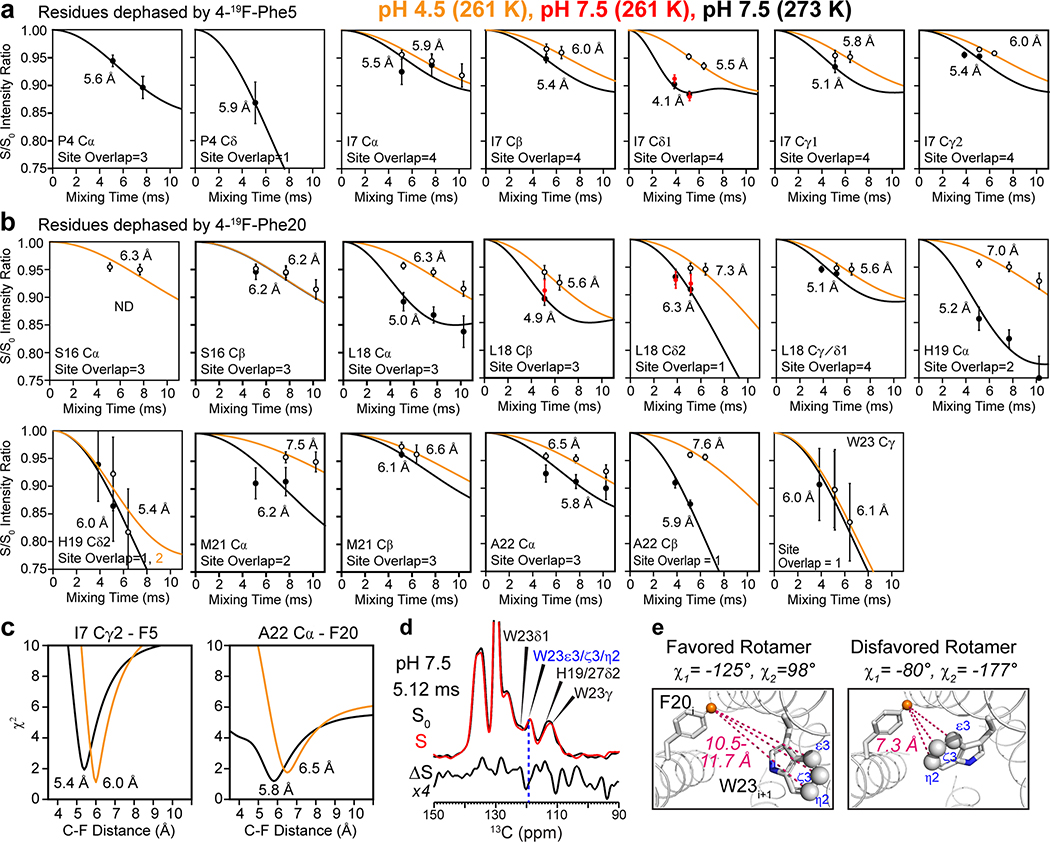
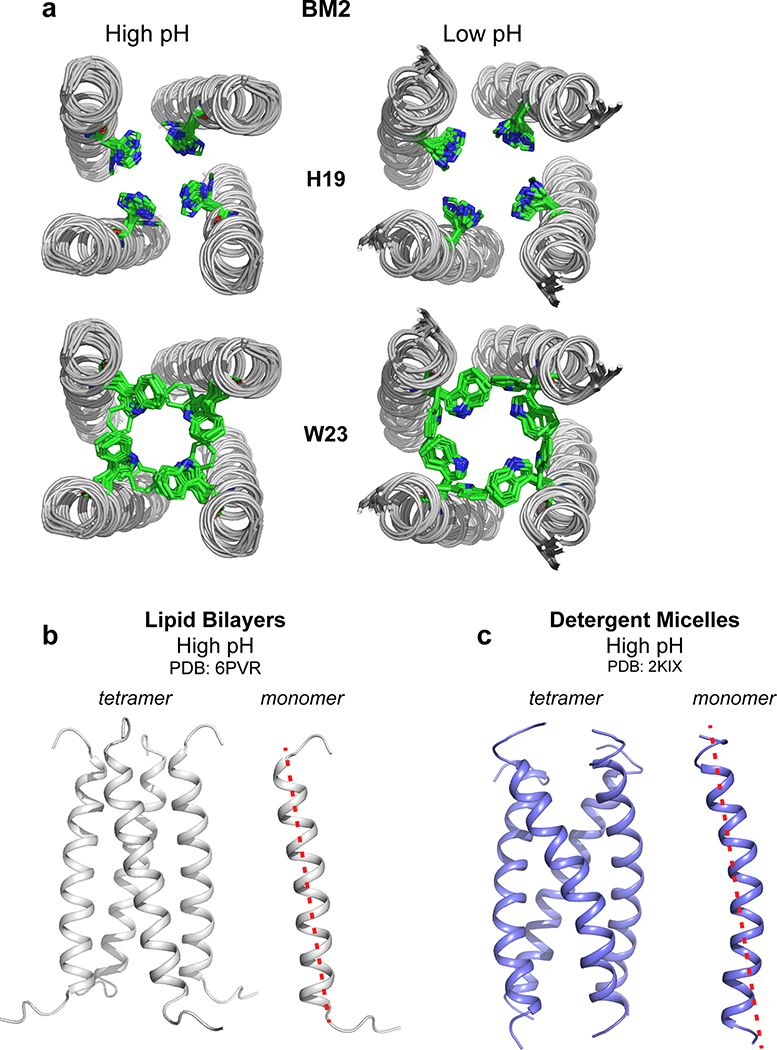
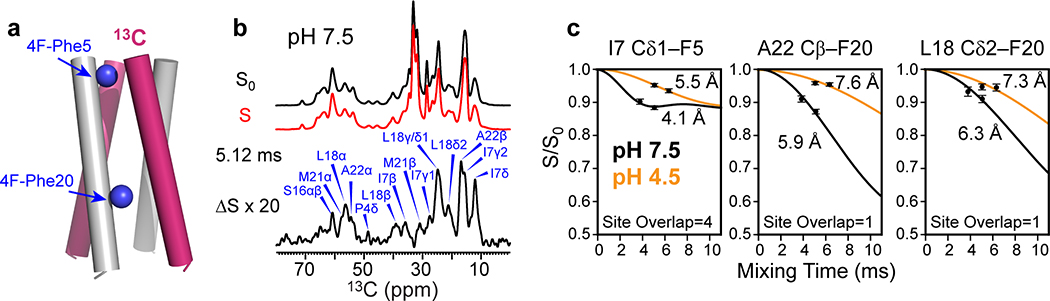
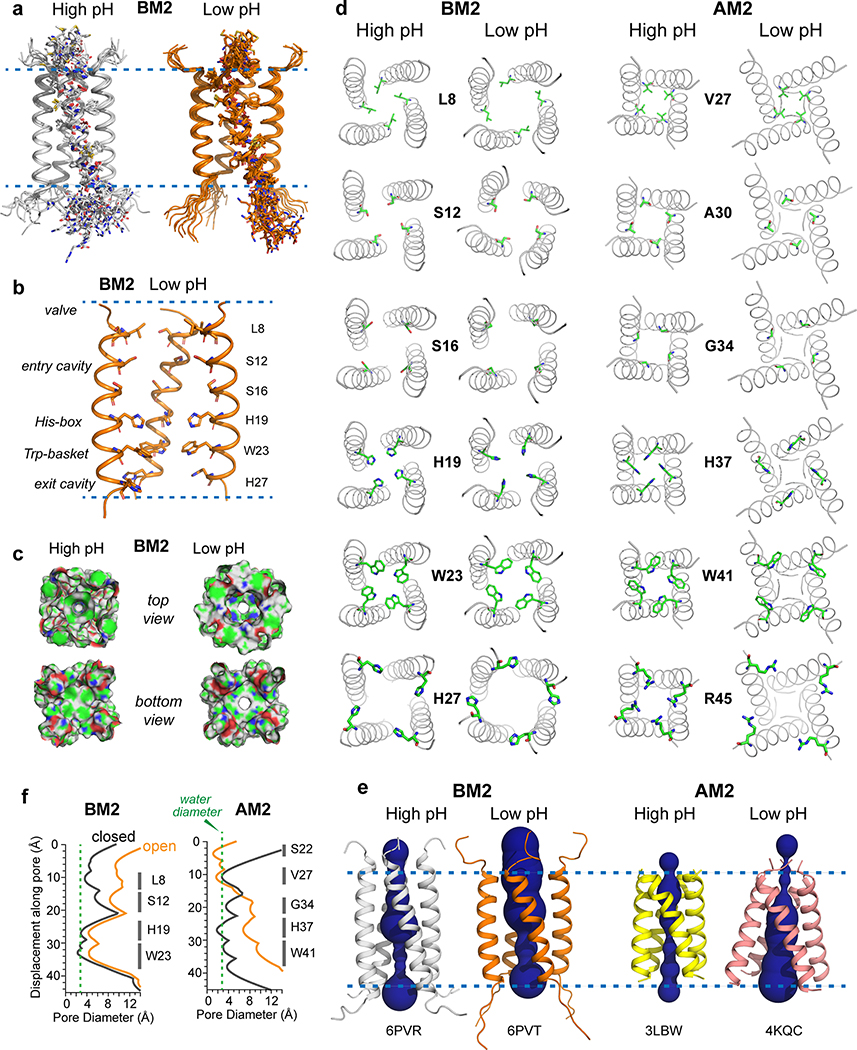
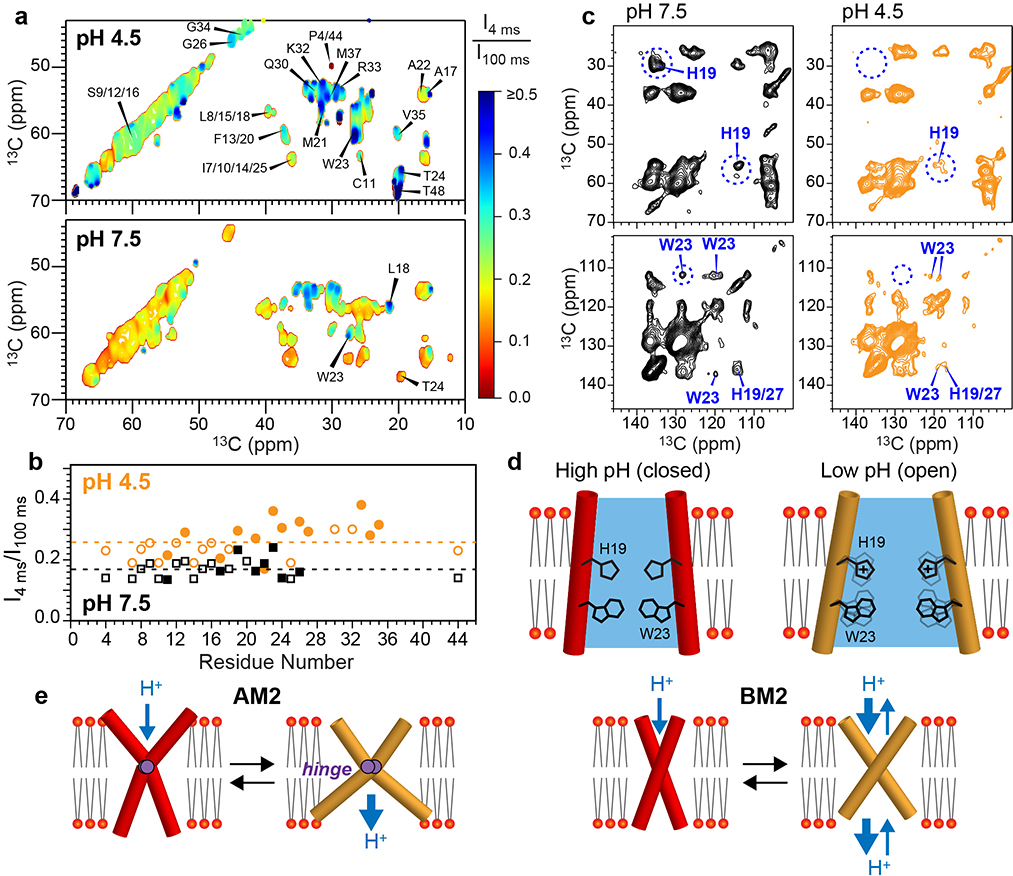
The influenza B M2 (BM2) proton channel is activated by acidic pH to mediate virus uncoating. Unlike influenza A M2 (AM2), which conducts protons with strong inward rectification, BM2 conducts protons both inward and outward. Here we report 1.4- and 1.5-Å solid-state NMR structures of the transmembrane domain of the closed and open BM2 channels in a phospholipid environment. Upon activation, the transmembrane helices increase the tilt angle by 6° and the average pore diameter enlarges by 2.1 Å. BM2 thus undergoes a scissor motion for activation, which differs from the alternating-access motion of AM2. These results indicate that asymmetric proton conduction requires a backbone hinge motion, whereas bidirectional conduction is achieved by a symmetric scissor motion. The proton-selective histidine and gating tryptophan in the open BM2 reorient on the microsecond timescale, similar to AM2, indicating that side chain dynamics are the essential driver of proton shuttling.
Conflict of interest statement
Competing interests
The authors declare no competing financial interests.
Figures
Similar articles
-
Structure and dynamics of the proton-selective histidine and the gating tryptophan in an inward rectifying hybrid influenza B and A virus M2 proton channel.Phys Chem Chem Phys. 2024 Jul 31;26(30):20629-20644. doi: 10.1039/d4cp01648c. Phys Chem Chem Phys. 2024. PMID: 39037444 Free PMC article.
-
Side Chain Structures of the Proton-Selective Histidine and Gating Tryptophan in Influenza BM2 Reveal Both Conservation and Variation of the Proton Conduction Mechanism.Biochemistry. 2025 Jul 15;64(14):3081-3092. doi: 10.1021/acs.biochem.5c00242. Epub 2025 Jun 24. Biochemistry. 2025. PMID: 40554716
-
Solid-State NMR Investigation of the Conformation, Proton Conduction, and Hydration of the Influenza B Virus M2 Transmembrane Proton Channel.J Am Chem Soc. 2016 Jul 6;138(26):8143-55. doi: 10.1021/jacs.6b03142. Epub 2016 Jun 23. J Am Chem Soc. 2016. PMID: 27286559 Free PMC article.
-
Flu channel drug resistance: a tale of two sites.Protein Cell. 2010 Mar;1(3):246-58. doi: 10.1007/s13238-010-0025-y. Epub 2010 Feb 23. Protein Cell. 2010. PMID: 21203971 Free PMC article. Review.
-
The Structure, Function, and Pathobiology of the Influenza A and B Virus Ion Channels.Cold Spring Harb Perspect Med. 2020 Nov 2;10(11):a038505. doi: 10.1101/cshperspect.a038505. Cold Spring Harb Perspect Med. 2020. PMID: 31988204 Free PMC article. Review.
Cited by
-
Characterization of the SARS-CoV-2 E Protein: Sequence, Structure, Viroporin, and Inhibitors.Protein Sci. 2021 Jun;30(6):1114-1130. doi: 10.1002/pro.4075. Epub 2021 Apr 13. Protein Sci. 2021. PMID: 33813796 Free PMC article. Review.
-
Solid-State NMR of Virus Membrane Proteins.Acc Chem Res. 2025 Mar 18;58(6):847-860. doi: 10.1021/acs.accounts.4c00800. Epub 2025 Feb 28. Acc Chem Res. 2025. PMID: 40019485 Free PMC article.
-
Virus Structures and Dynamics by Magic-Angle Spinning NMR.Annu Rev Virol. 2021 Sep 29;8(1):219-237. doi: 10.1146/annurev-virology-011921-064653. Annu Rev Virol. 2021. PMID: 34586870 Free PMC article. Review.
-
Structure and drug binding of the SARS-CoV-2 envelope protein transmembrane domain in lipid bilayers.Nat Struct Mol Biol. 2020 Dec;27(12):1202-1208. doi: 10.1038/s41594-020-00536-8. Epub 2020 Nov 11. Nat Struct Mol Biol. 2020. PMID: 33177698 Free PMC article.
-
Functions of Viroporins in the Viral Life Cycle and Their Regulation of Host Cell Responses.Front Immunol. 2022 Jun 2;13:890549. doi: 10.3389/fimmu.2022.890549. eCollection 2022. Front Immunol. 2022. PMID: 35720341 Free PMC article. Review.
References
-
- Mitchell P A general theory of membrane transport from studies of bacteria. Nature 180, 134–136 (1957). - PubMed
-
- Forrest LR, Krämer R & Ziegler C The structural basis of secondary active transport mechanisms. Biochim. Biophys. Acta 1807, 167–188 (2011). - PubMed
-
- Pinto LH & Lamb RA The M2 proton channels of influenza A and B viruses. J. Biol. Chem 281, 8997–9000 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
