

The role of cell lineage in the development of neuronal circuitry and function
- PMID: 32017903
- PMCID: PMC7394725
- DOI: 10.1016/j.ydbio.2020.01.012
The role of cell lineage in the development of neuronal circuitry and function
Abstract
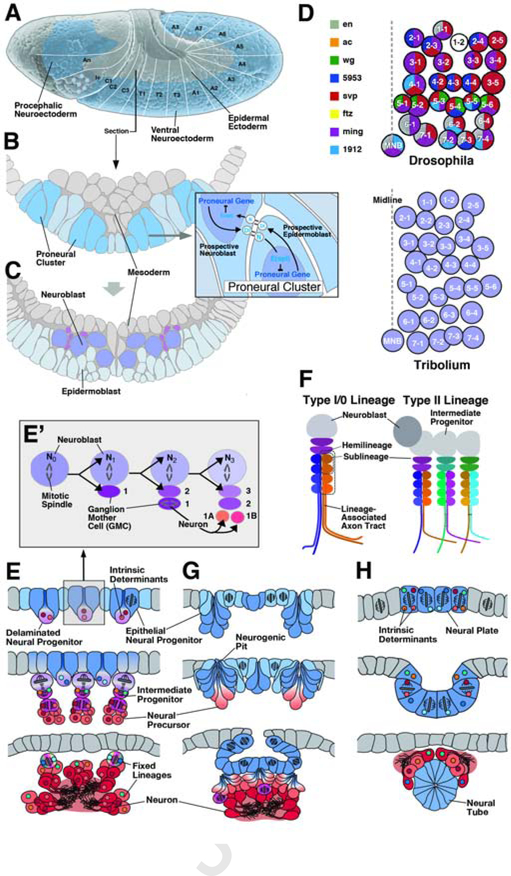
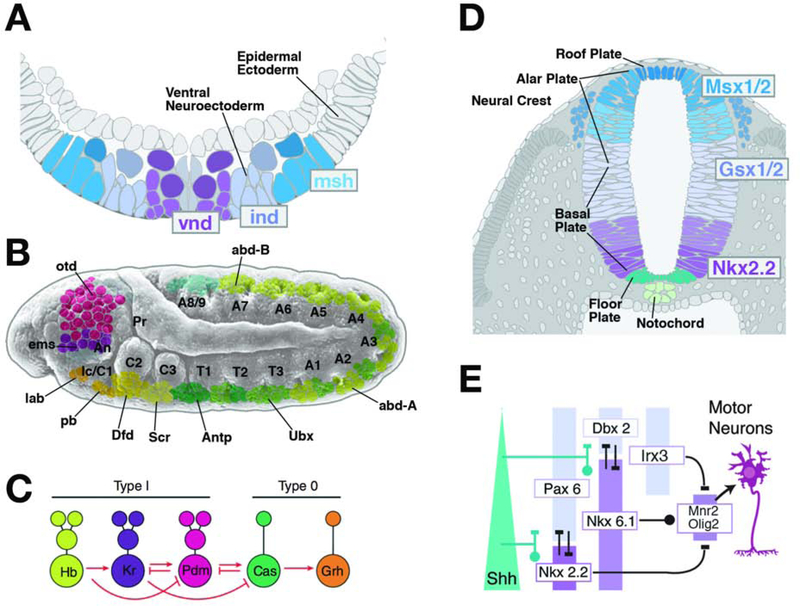
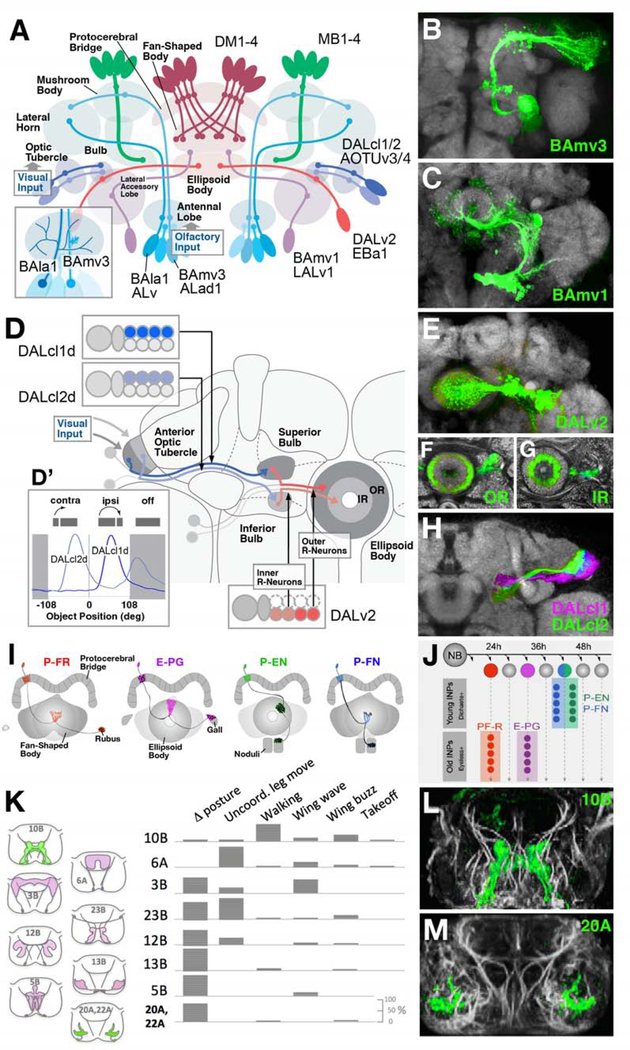
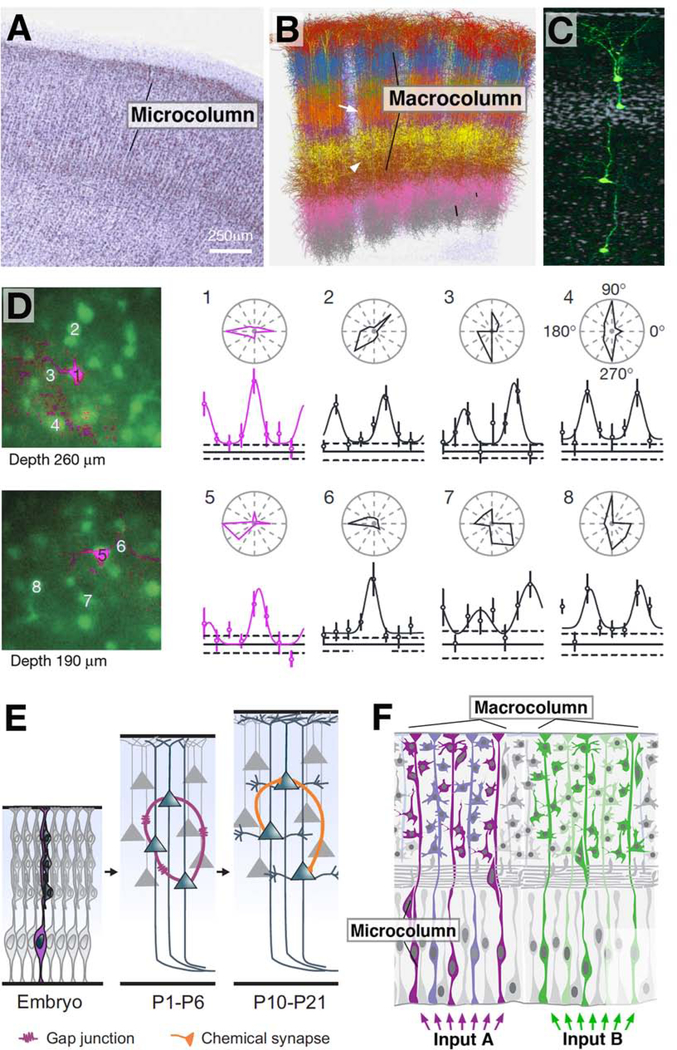
Complex nervous systems have a modular architecture, whereby reiterative groups of neurons ("modules") that share certain structural and functional properties are integrated into large neural circuits. Neurons develop from proliferating progenitor cells that, based on their location and time of appearance, are defined by certain genetic programs. Given that genes expressed by a given progenitor play a fundamental role in determining the properties of its lineage (i.e., the neurons descended from that progenitor), one efficient developmental strategy would be to have lineages give rise to the structural modules of the mature nervous system. It is clear that this strategy plays an important role in neural development of many invertebrate animals, notably insects, where the availability of genetic techniques has made it possible to analyze the precise relationship between neuronal origin and differentiation since several decades. Similar techniques, developed more recently in the vertebrate field, reveal that functional modules of the mammalian cerebral cortex are also likely products of developmentally defined lineages. We will review studies that relate cell lineage to circuitry and function from a comparative developmental perspective, aiming at enhancing our understanding of neural progenitors and their lineages, and translating findings acquired in different model systems into a common conceptual framework.
Keywords: Cell fate; Cell lineage; Circuitry; Drosophila; Mouse; Neural progenitor; Proliferation; Xenopus; Zebrafish.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures

References
-
- Akam M, 1987. The molecular basis for metameric pattern in the Drosophila embryo. Development 101,1–22. - PubMed
-
- Alexandre P, Reugels AM, Barker D, Blanc E, Clarke JD, 2010.Neurons derive from the more apical daughter in asymmetric divisions in the zebrafish neural tube. Nat. Neurosci. 13, 673–9. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
