

The Non-Coding RNA Landscape of Plasma Cell Dyscrasias
- PMID: 32019064
- PMCID: PMC7072200
- DOI: 10.3390/cancers12020320
The Non-Coding RNA Landscape of Plasma Cell Dyscrasias
Abstract
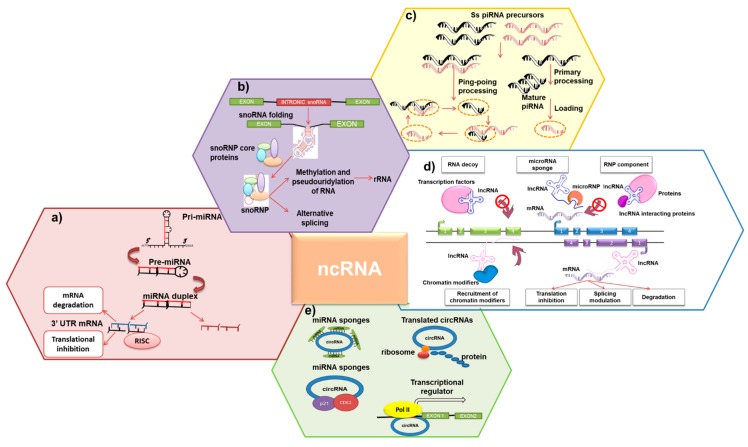
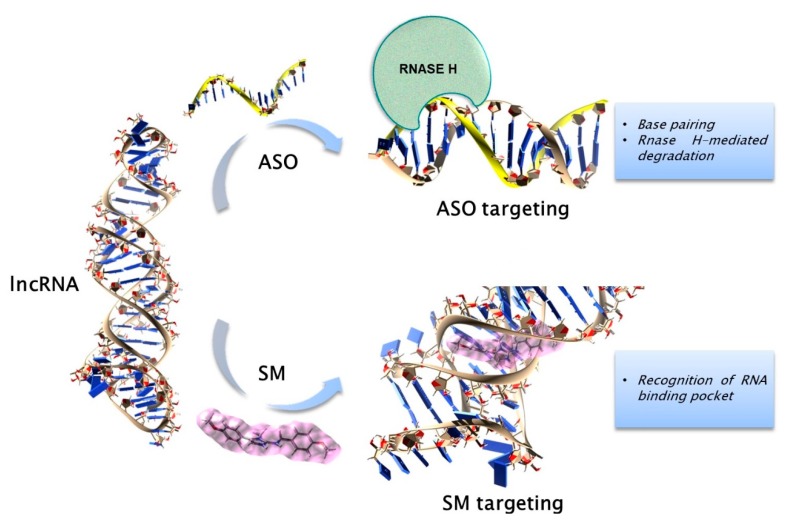
Despite substantial advancements have been done in the understanding of the pathogenesis of plasma cell (PC) disorders, these malignancies remain hard-to-treat. The discovery and subsequent characterization of non-coding transcripts, which include several members with diverse length and mode of action, has unraveled novel mechanisms of gene expression regulation often malfunctioning in cancer. Increasing evidence indicates that such non-coding molecules also feature in the pathobiology of PC dyscrasias, where they are endowed with strong therapeutic and/or prognostic potential. In this review, we aim to summarize the most relevant findings on the biological and clinical features of the non-coding RNA landscape of malignant PCs, with major focus on multiple myeloma. The most relevant classes of non-coding RNAs will be examined, along with the mechanisms accounting for their dysregulation and the recent strategies used for their targeting in PC dyscrasias. It is hoped these insights may lead to clinical applications of non-coding RNA molecules as biomarkers or therapeutic targets/agents in the near future.
Keywords: lncRNA; miRNA; multiple myeloma; non-coding RNA; plasma cell dyscrasia.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- Beason T.S., Chang S.H., Sanfilippo K.M., Luo S., Colditz G.A., Vij R., Tomasson M.H., Dipersio J.F., Stockerl-Goldstein K., Ganti A., et al. Influence of body mass index on survival in veterans with multiple myeloma. Oncologist. 2013;18:1074–1079. doi: 10.1634/theoncologist.2013-0015. - DOI - PMC - PubMed
