

Polyglutamine-Expanded Androgen Receptor Alteration of Skeletal Muscle Homeostasis and Myonuclear Aggregation Are Affected by Sex, Age and Muscle Metabolism
- PMID: 32019272
- PMCID: PMC7072234
- DOI: 10.3390/cells9020325
Polyglutamine-Expanded Androgen Receptor Alteration of Skeletal Muscle Homeostasis and Myonuclear Aggregation Are Affected by Sex, Age and Muscle Metabolism
Abstract
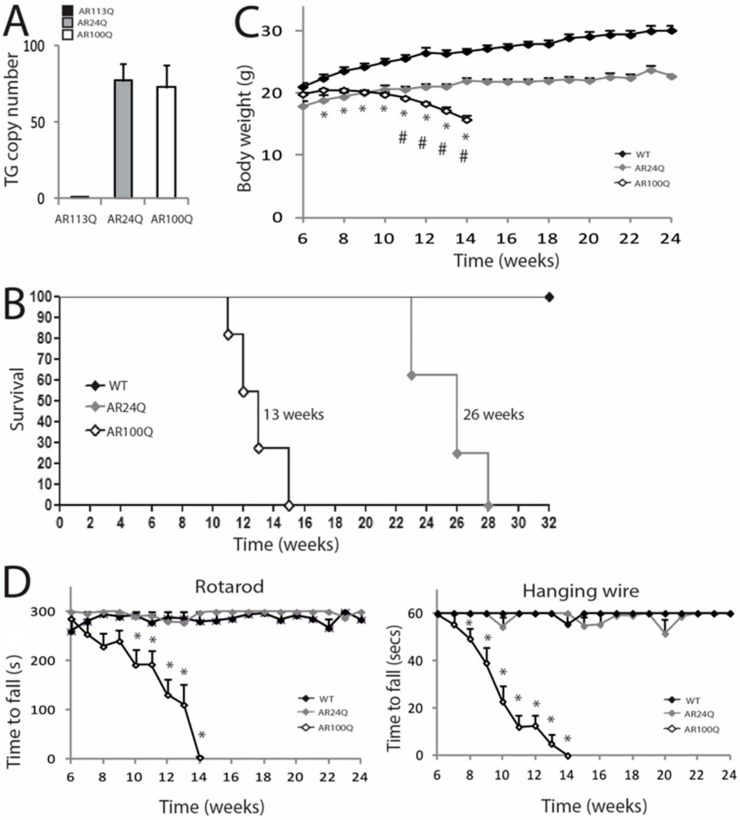
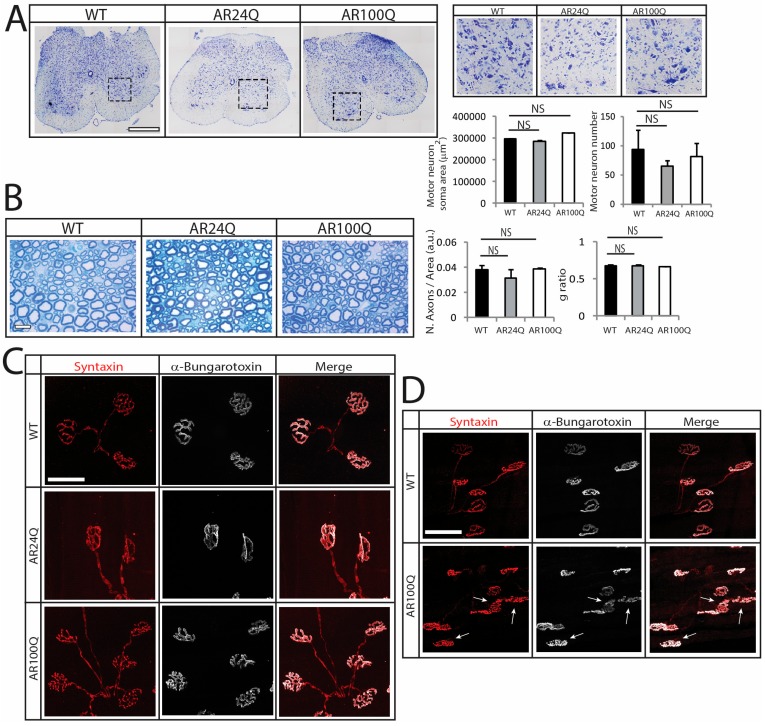
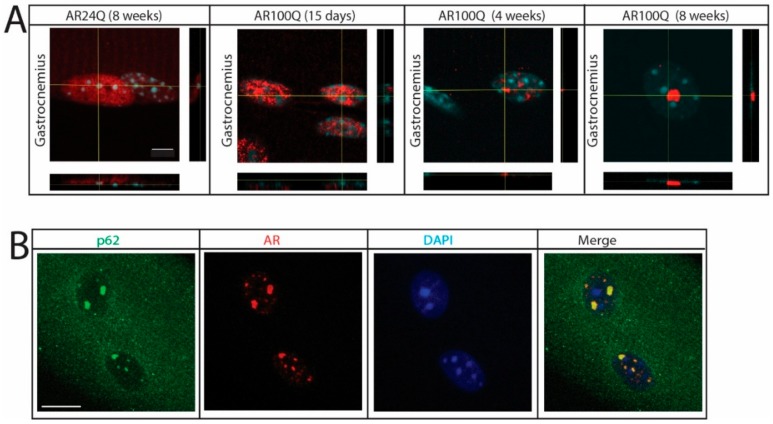
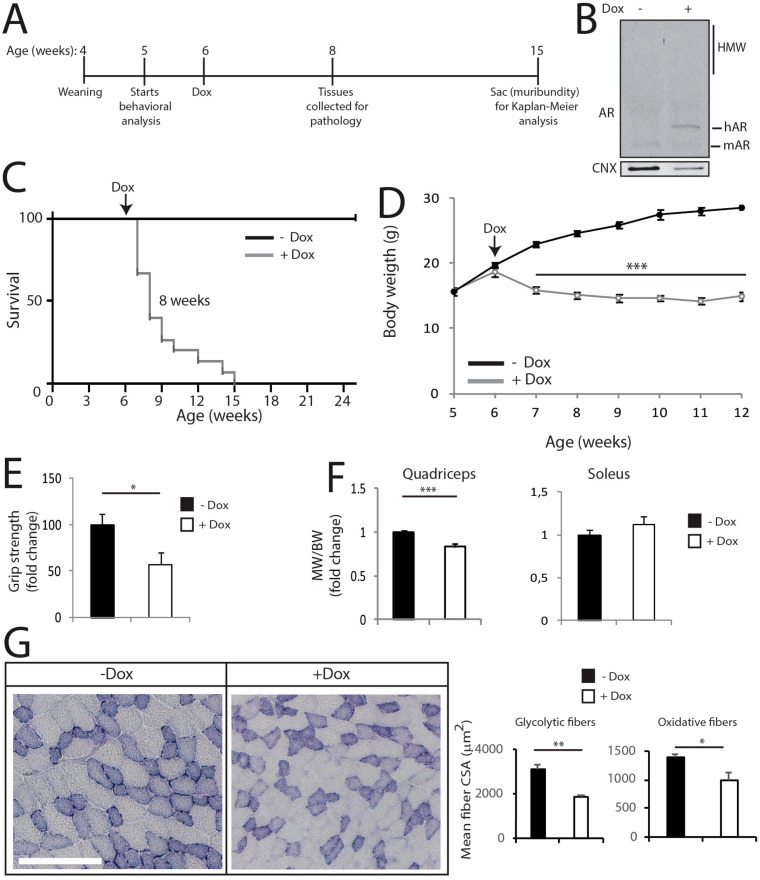
Polyglutamine (polyQ) expansions in the androgen receptor (AR) gene cause spinal and bulbar muscular atrophy (SBMA), a neuromuscular disease characterized by lower motor neuron (MN) loss and skeletal muscle atrophy, with an unknown mechanism. We generated new mouse models of SBMA for constitutive and inducible expression of mutant AR and performed biochemical, histological and functional analyses of phenotype. We show that polyQ-expanded AR causes motor dysfunction, premature death, IIb-to-IIa/IIx fiber-type change, glycolytic-to-oxidative fiber-type switching, upregulation of atrogenes and autophagy genes and mitochondrial dysfunction in skeletal muscle, together with signs of muscle denervation at late stage of disease. PolyQ expansions in the AR resulted in nuclear enrichment. Within the nucleus, mutant AR formed 2% sodium dodecyl sulfate (SDS)-resistant aggregates and inclusion bodies in myofibers, but not spinal cord and brainstem, in a process exacerbated by age and sex. Finally, we found that two-week induction of expression of polyQ-expanded AR in adult mice was sufficient to cause premature death, body weight loss and muscle atrophy, but not aggregation, metabolic alterations, motor coordination and fiber-type switch, indicating that expression of the disease protein in the adulthood is sufficient to recapitulate several, but not all SBMA manifestations in mice. These results imply that chronic expression of polyQ-expanded AR, i.e. during development and prepuberty, is key to induce the full SBMA muscle pathology observed in patients. Our data support a model whereby chronic expression of polyQ-expanded AR triggers muscle atrophy through toxic (neomorphic) gain of function mechanisms distinct from normal (hypermorphic) gain of function mechanisms.
Keywords: 2% SDS-resistant aggregates; androgen receptor; inclusion bodies; muscle metabolism; polyglutamine diseases; skeletal muscle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
