

Deciphering the Rules Underlying Xenogeneic Silencing and Counter-Silencing of Lsr2-like Proteins Using CgpS of Corynebacterium glutamicum as a Model
- PMID: 32019787
- PMCID: PMC7002338
- DOI: 10.1128/mBio.02273-19
Deciphering the Rules Underlying Xenogeneic Silencing and Counter-Silencing of Lsr2-like Proteins Using CgpS of Corynebacterium glutamicum as a Model
Abstract
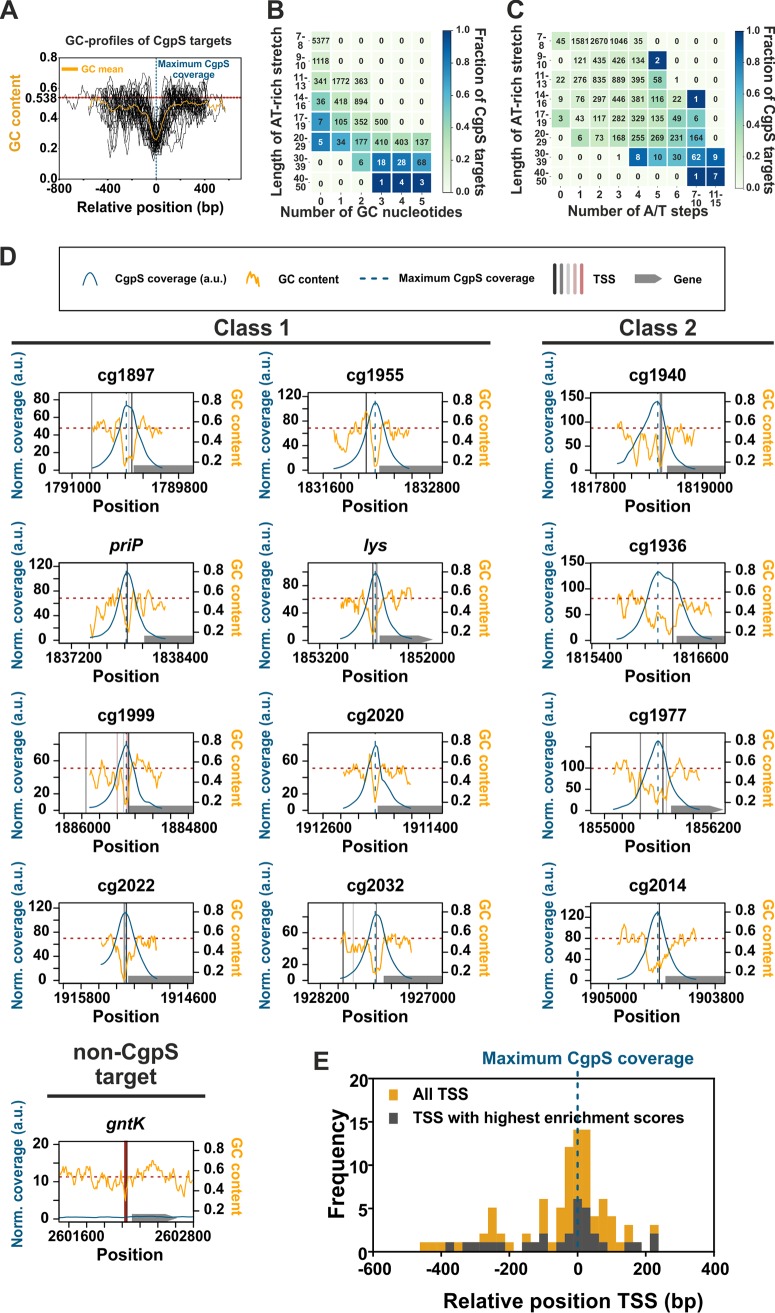
Lsr2-like nucleoid-associated proteins play an important role as xenogeneic silencers (XS) of horizontally acquired genomic regions in actinobacteria. In this study, we systematically analyzed the in vivo constraints underlying silencing and counter-silencing of the Lsr2-like protein CgpS in Corynebacterium glutamicum Genome-wide analysis revealed binding of CgpS to regions featuring a distinct drop in GC profile close to the transcription start site (TSS) but also identified an overrepresented motif with multiple A/T steps at the nucleation site of the nucleoprotein complex. Binding of specific transcription factors (TFs) may oppose XS activity, leading to counter-silencing. Following a synthetic counter-silencing approach, target gene activation was realized by inserting operator sites of an effector-responsive TF within various CgpS target promoters, resulting in increased promoter activity upon TF binding. Analysis of reporter constructs revealed maximal counter-silencing when the TF operator site was inserted at the position of maximal CgpS coverage. This principle was implemented in a synthetic toggle switch, which features a robust and reversible response to effector availability, highlighting the potential for biotechnological applications. Together, our results provide comprehensive insights into how Lsr2 silencing and counter-silencing shape evolutionary network expansion in this medically and biotechnologically relevant bacterial phylum.IMPORTANCE In actinobacteria, Lsr2-like nucleoid-associated proteins function as xenogeneic silencers (XS) of horizontally acquired genomic regions, including viral elements, virulence gene clusters in Mycobacterium tuberculosis, and genes involved in cryptic specialized metabolism in Streptomyces species. Consequently, a detailed mechanistic understanding of Lsr2 binding in vivo is relevant as a potential drug target and for the identification of novel bioactive compounds. Here, we followed an in vivo approach to investigate the rules underlying xenogeneic silencing and counter-silencing of the Lsr2-like XS CgpS from Corynebacterium glutamicum Our results demonstrated that CgpS distinguishes between self and foreign by recognizing a distinct drop in GC profile in combination with a short, sequence-specific motif at the nucleation site. Following a synthetic counter-silencer approach, we studied the potential and constraints of transcription factors to counteract CgpS silencing, thereby facilitating the integration of new genetic traits into host regulatory networks.
Keywords: AT-rich DNA; Lsr2; actinobacteria; counter-silencing; horizontal gene transfer; regulatory networks; xenogeneic silencing.
Copyright © 2020 Wiechert et al.
Figures





Similar articles
-
CRISPR/dCas-mediated counter-silencing: reprogramming dCas proteins into antagonists of xenogeneic silencers.mBio. 2025 Jul 9;16(7):e0038225. doi: 10.1128/mbio.00382-25. Epub 2025 May 28. mBio. 2025. PMID: 40434115 Free PMC article.
-
Inducible Expression Systems Based on Xenogeneic Silencing and Counter-Silencing and Design of a Metabolic Toggle Switch.ACS Synth Biol. 2020 Aug 21;9(8):2023-2038. doi: 10.1021/acssynbio.0c00111. Epub 2020 Jul 27. ACS Synth Biol. 2020. PMID: 32649183 Free PMC article.
-
Silencing of cryptic prophages in Corynebacterium glutamicum.Nucleic Acids Res. 2016 Dec 1;44(21):10117-10131. doi: 10.1093/nar/gkw692. Epub 2016 Aug 4. Nucleic Acids Res. 2016. PMID: 27492287 Free PMC article.
-
Xenogeneic Silencing and Bacterial Genome Evolution: Mechanisms for DNA Recognition Imply Multifaceted Roles of Xenogeneic Silencers.Mol Biol Evol. 2021 Sep 27;38(10):4135-4148. doi: 10.1093/molbev/msab136. Mol Biol Evol. 2021. PMID: 34003286 Free PMC article. Review.
-
Impact of Xenogeneic Silencing on Phage-Host Interactions.J Mol Biol. 2019 Nov 22;431(23):4670-4683. doi: 10.1016/j.jmb.2019.02.011. Epub 2019 Feb 21. J Mol Biol. 2019. PMID: 30796986 Free PMC article. Review.
Cited by
-
A nucleoid-associated protein is involved in the emergence of antibiotic resistance by promoting the frequent exchange of the replicative DNA polymerase in M. smegmatis.bioRxiv [Preprint]. 2024 Jan 3:2023.06.12.544663. doi: 10.1101/2023.06.12.544663. bioRxiv. 2024. Update in: mSphere. 2024 May 29;9(5):e0012224. doi: 10.1128/msphere.00122-24. PMID: 38260554 Free PMC article. Updated. Preprint.
-
Phylogenetic Distribution of WhiB- and Lsr2-Type Regulators in Actinobacteriophage Genomes.Microbiol Spectr. 2021 Dec 22;9(3):e0072721. doi: 10.1128/Spectrum.00727-21. Epub 2021 Nov 24. Microbiol Spectr. 2021. PMID: 34817283 Free PMC article.
-
Genome Sequence of the Bacteriophage CL31 and Interaction with the Host Strain Corynebacterium glutamicum ATCC 13032.Viruses. 2021 Mar 17;13(3):495. doi: 10.3390/v13030495. Viruses. 2021. PMID: 33802915 Free PMC article.
-
A nucleoid-associated protein is involved in the emergence of antibiotic resistance by promoting the frequent exchange of the replicative DNA polymerase in Mycobacterium smegmatis.mSphere. 2024 May 29;9(5):e0012224. doi: 10.1128/msphere.00122-24. Epub 2024 Apr 9. mSphere. 2024. PMID: 38591887 Free PMC article.
-
CRISPR/dCas-mediated counter-silencing: reprogramming dCas proteins into antagonists of xenogeneic silencers.mBio. 2025 Jul 9;16(7):e0038225. doi: 10.1128/mbio.00382-25. Epub 2025 May 28. mBio. 2025. PMID: 40434115 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
