

Pseudomonas aeruginosa exoenzyme Y directly bundles actin filaments
- PMID: 32019868
- PMCID: PMC7076225
- DOI: 10.1074/jbc.RA119.012320
Pseudomonas aeruginosa exoenzyme Y directly bundles actin filaments
Abstract
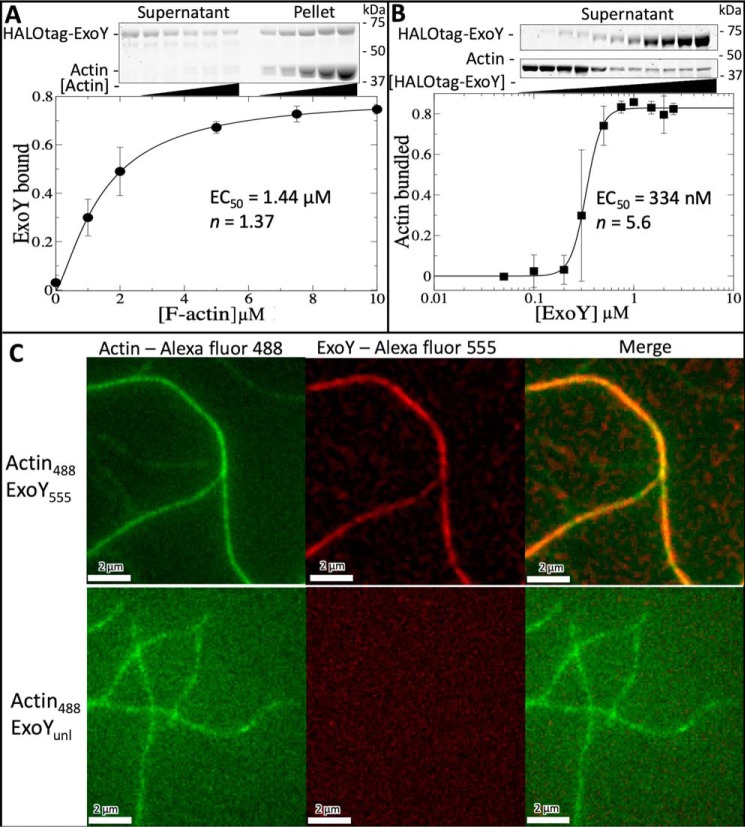
Pseudomonas aeruginosa uses a type III secretion system (T3SS) to inject cytotoxic effector proteins into host cells. The promiscuous nucleotidyl cyclase, exoenzyme Y (ExoY), is one of the most common effectors found in clinical P. aeruginosa isolates. Recent studies have revealed that the nucleotidyl cyclase activity of ExoY is stimulated by actin filaments (F-actin) and that ExoY alters actin cytoskeleton dynamics in vitro, via an unknown mechanism. The actin cytoskeleton plays an important role in numerous key biological processes and is targeted by many pathogens to gain competitive advantages. We utilized total internal reflection fluorescence microscopy, bulk actin assays, and EM to investigate how ExoY impacts actin dynamics. We found that ExoY can directly bundle actin filaments with high affinity, comparable with eukaryotic F-actin-bundling proteins, such as fimbrin. Of note, ExoY enzymatic activity was not required for F-actin bundling. Bundling is known to require multiple actin-binding sites, yet small-angle X-ray scattering experiments revealed that ExoY is a monomer in solution, and previous data suggested that ExoY possesses only one actin-binding site. We therefore hypothesized that ExoY oligomerizes in response to F-actin binding and have used the ExoY structure to construct a dimer-based structural model for the ExoY-F-actin complex. Subsequent mutational analyses suggested that the ExoY oligomerization interface plays a crucial role in mediating F-actin bundling. Our results indicate that ExoY represents a new class of actin-binding proteins that modulate the actin cytoskeleton both directly, via F-actin bundling, and indirectly, via actin-activated nucleotidyl cyclase activity.
Keywords: Pseudomonas aeruginosa; actin; actin bundling; bacterial pathogenesis; bacterial toxin; cytoskeleton; exoenzyme Y (ExoY); host-microbe interactions; host-pathogen interaction; protein-protein interaction; virulence factor.
© 2020 Mancl et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures





References
-
- Micek S. T., Wunderink R. G., Kollef M. H., Chen C., Rello J., Chastre J., Antonelli M., Welte T., Clair B., Ostermann H., Calbo E., Torres A., Menichetti F., Schramm G. E., and Menon V. (2015) An international multicenter retrospective study of Pseudomonas aeruginosa nosocomial pneumonia: impact of multidrug resistance. Crit. Care 19, 219 10.1186/s13054-015-0926-5 - DOI - PMC - PubMed
-
- Sato H., Frank D. W., Hillard C. J., Feix J. B., Pankhaniya R. R., Moriyama K., Finck-Barbançon V., Buchaklian A., Lei M., Long R. M., Wiener-Kronish J., and Sawa T. (2003) The mechanism of action of the Pseudomonas aeruginosa-encoded type III cytotoxin, ExoU. EMBO J. 22, 2959–2969 10.1093/emboj/cdg290 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
