

Control of XPR1-dependent cellular phosphate efflux by InsP8 is an exemplar for functionally-exclusive inositol pyrophosphate signaling
- PMID: 32019887
- PMCID: PMC7035621
- DOI: 10.1073/pnas.1908830117
Control of XPR1-dependent cellular phosphate efflux by InsP8 is an exemplar for functionally-exclusive inositol pyrophosphate signaling
Abstract
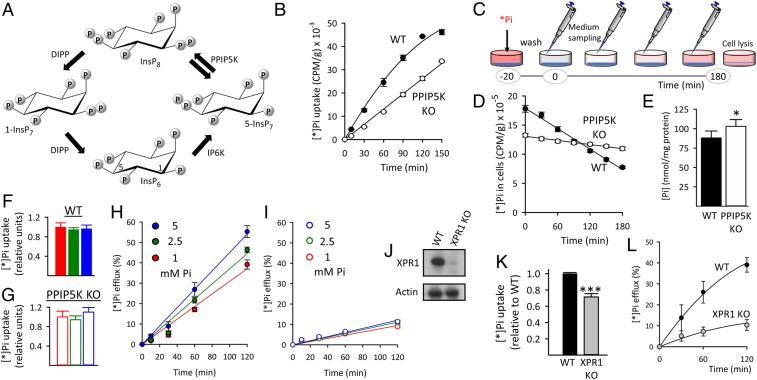
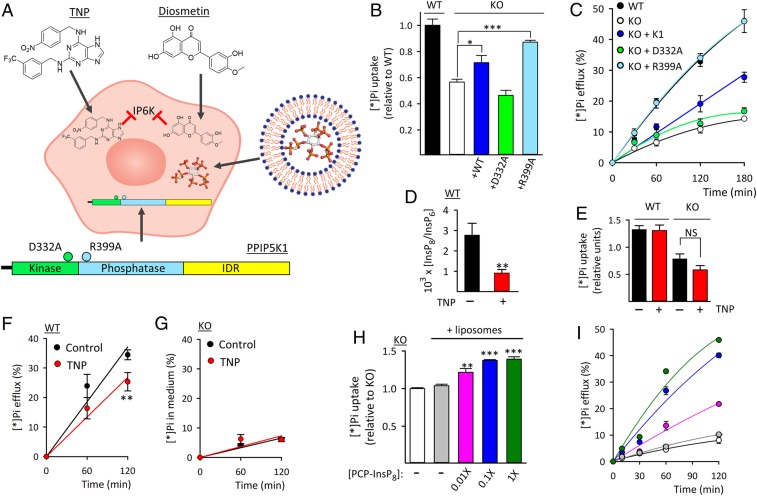
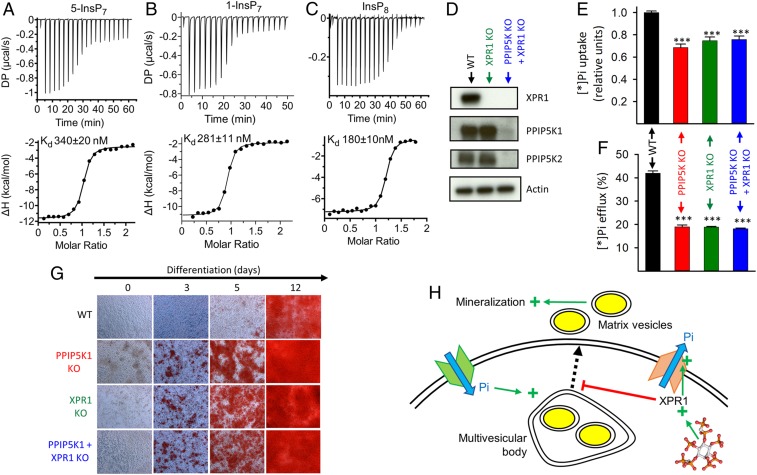
Homeostasis of cellular fluxes of inorganic phosphate (Pi) supervises its structural roles in bones and teeth, its pervasive regulation of cellular metabolism, and its functionalization of numerous organic compounds. Cellular Pi efflux is heavily reliant on Xenotropic and Polytropic Retrovirus Receptor 1 (XPR1), regulation of which is largely unknown. We demonstrate specificity of XPR1 regulation by a comparatively uncharacterized member of the inositol pyrophosphate (PP-InsP) signaling family: 1,5-bis-diphosphoinositol 2,3,4,6-tetrakisphosphate (InsP8). XPR1-mediated Pi efflux was inhibited by reducing cellular InsP8 synthesis, either genetically (knockout [KO] of diphosphoinositol pentakisphosphate kinases [PPIP5Ks] that synthesize InsP8) or pharmacologically [cell treatment with 2.5 µM dietary flavonoid or 10 µM N2-(m-trifluorobenzyl), N6-(p-nitrobenzyl) purine], to inhibit inositol hexakisphosphate kinases upstream of PPIP5Ks. Attenuated Pi efflux from PPIP5K KO cells was quantitatively phenocopied by KO of XPR1 itself. Moreover, Pi efflux from PPIP5K KO cells was rescued by restoration of InsP8 levels through transfection of wild-type PPIP5K1; transfection of kinase-dead PPIP5K1 was ineffective. Pi efflux was also rescued in a dose-dependent manner by liposomal delivery of a metabolically resistant methylene bisphosphonate (PCP) analog of InsP8; PCP analogs of other PP-InsP signaling molecules were ineffective. High-affinity binding of InsP8 to the XPR1 N-terminus (Kd = 180 nM) was demonstrated by isothermal titration calorimetry. To derive a cellular biology perspective, we studied biomineralization in the Soas-2 osteosarcoma cell line. KO of PPIP5Ks or XPR1 strongly reduced Pi efflux and accelerated differentiation to the mineralization end point. We propose that catalytically compromising PPIP5K mutations might extend an epistatic repertoire for XPR1 dysregulation, with pathological consequences for bone maintenance and ectopic calcification.
Keywords: inositol; metabolism; signaling.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Interplay between primary familial brain calcification-associated SLC20A2 and XPR1 phosphate transporters requires inositol polyphosphates for control of cellular phosphate homeostasis.J Biol Chem. 2020 Jul 10;295(28):9366-9378. doi: 10.1074/jbc.RA119.011376. Epub 2020 May 11. J Biol Chem. 2020. PMID: 32393577 Free PMC article.
-
KO of 5-InsP7 kinase activity transforms the HCT116 colon cancer cell line into a hypermetabolic, growth-inhibited phenotype.Proc Natl Acad Sci U S A. 2017 Nov 7;114(45):11968-11973. doi: 10.1073/pnas.1702370114. Epub 2017 Oct 25. Proc Natl Acad Sci U S A. 2017. PMID: 29078269 Free PMC article.
-
The Significance of the Bifunctional Kinase/Phosphatase Activities of Diphosphoinositol Pentakisphosphate Kinases (PPIP5Ks) for Coupling Inositol Pyrophosphate Cell Signaling to Cellular Phosphate Homeostasis.J Biol Chem. 2017 Mar 17;292(11):4544-4555. doi: 10.1074/jbc.M116.765743. Epub 2017 Jan 26. J Biol Chem. 2017. PMID: 28126903 Free PMC article.
-
Inositol Pyrophosphate Pathways and Mechanisms: What Can We Learn from Plants?Molecules. 2020 Jun 17;25(12):2789. doi: 10.3390/molecules25122789. Molecules. 2020. PMID: 32560343 Free PMC article. Review.
-
Role of transporters in regulating mammalian intracellular inorganic phosphate.Front Pharmacol. 2023 Mar 30;14:1163442. doi: 10.3389/fphar.2023.1163442. eCollection 2023. Front Pharmacol. 2023. PMID: 37063296 Free PMC article. Review.
Cited by
-
Encapsulation of Inositol Hexakisphosphate with Chitosan via Gelation to Facilitate Cellular Delivery and Programmed Cell Death in Human Breast Cancer Cells.Bioengineering (Basel). 2024 Sep 17;11(9):931. doi: 10.3390/bioengineering11090931. Bioengineering (Basel). 2024. PMID: 39329673 Free PMC article.
-
InsP7 is a small-molecule regulator of NUDT3-mediated mRNA decapping and processing-body dynamics.Proc Natl Acad Sci U S A. 2020 Aug 11;117(32):19245-19253. doi: 10.1073/pnas.1922284117. Epub 2020 Jul 29. Proc Natl Acad Sci U S A. 2020. PMID: 32727897 Free PMC article.
-
The Solute Carrier (SLC) Transporter Superfamily as Therapeutic Targets for the Treatment of Head and Neck Squamous Cell Carcinoma.Cancers (Basel). 2024 Sep 22;16(18):3226. doi: 10.3390/cancers16183226. Cancers (Basel). 2024. PMID: 39335197 Free PMC article.
-
The Role of Inositol Hexakisphosphate Kinase in the Central Nervous System.Biomolecules. 2023 Aug 28;13(9):1317. doi: 10.3390/biom13091317. Biomolecules. 2023. PMID: 37759717 Free PMC article. Review.
-
A genomic estimated breeding value-assisted reduction method of single nucleotide polymorphism sets: a novel approach for determining the cutoff thresholds in genome-wide association studies and best linear unbiased prediction.Anim Cells Syst (Seoul). 2023 Sep 2;27(1):180-186. doi: 10.1080/19768354.2023.2250841. eCollection 2023. Anim Cells Syst (Seoul). 2023. PMID: 37674816 Free PMC article.
References
-
- Quintáns B., Oliveira J., Sobrido M. J., Primary familial brain calcifications. Handb. Clin. Neurol. 147, 307–317 (2018). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
