

Nucleosomal DNA Dynamics Mediate Oct4 Pioneer Factor Binding
- PMID: 32027821
- PMCID: PMC7202942
- DOI: 10.1016/j.bpj.2019.12.038
Nucleosomal DNA Dynamics Mediate Oct4 Pioneer Factor Binding
Abstract
Transcription factor (TF) proteins bind to DNA to regulate gene expression. Normally, accessibility to DNA is required for their function. However, in the nucleus, the DNA is often inaccessible, wrapped around histone proteins in nucleosomes forming the chromatin. Pioneer TFs are thought to induce chromatin opening by recognizing their DNA binding sites on nucleosomes. For example, Oct4, a master regulator and inducer of stem cell pluripotency, binds to DNA in nucleosomes in a sequence-specific manner. Here, we reveal the structural dynamics of nucleosomes that mediate Oct4 binding from molecular dynamics simulations. Nucleosome flexibility and the amplitude of nucleosome motions such as breathing and twisting are enhanced in nucleosomes with multiple TF binding sites. Moreover, the regions around the binding sites display higher local structural flexibility. Probing different structures of Oct4-nucleosome complexes, we show that alternative configurations in which Oct4 recognizes partial binding sites display stable TF-DNA interactions similar to those observed in complexes with free DNA and compatible with the DNA curvature and DNA-histone interactions. Therefore, we propose a structural basis for nucleosome recognition by a pioneer TF that is essential for understanding how chromatin is unraveled during cell fate conversions.
Copyright © 2020 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures

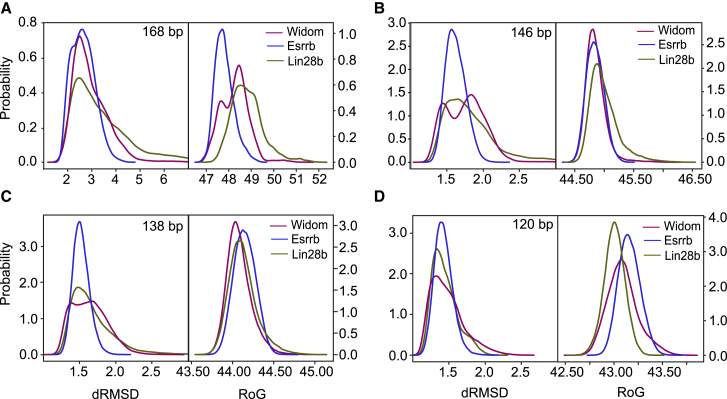
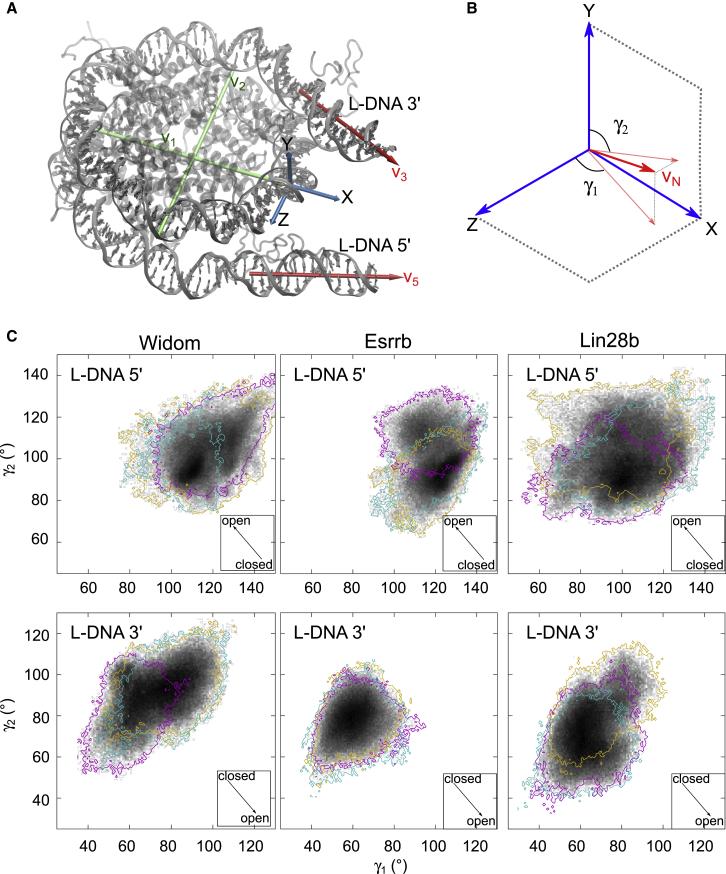



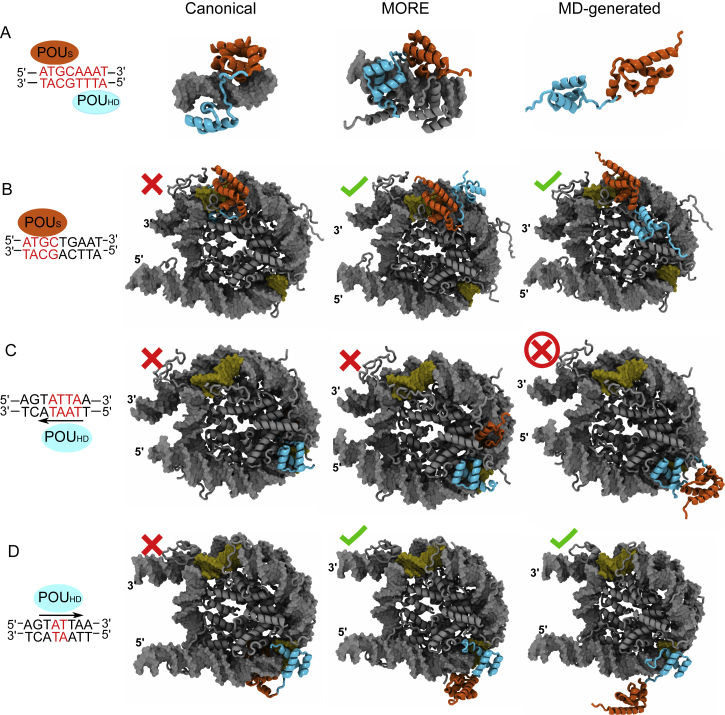
References
-
- Luger K., Mäder A.W., Richmond T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–260. - PubMed
-
- Takahashi K., Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
