

Optimizing Nervous System-Specific Gene Targeting with Cre Driver Lines: Prevalence of Germline Recombination and Influencing Factors
- PMID: 32027825
- PMCID: PMC7377387
- DOI: 10.1016/j.neuron.2020.01.008
Optimizing Nervous System-Specific Gene Targeting with Cre Driver Lines: Prevalence of Germline Recombination and Influencing Factors
Abstract
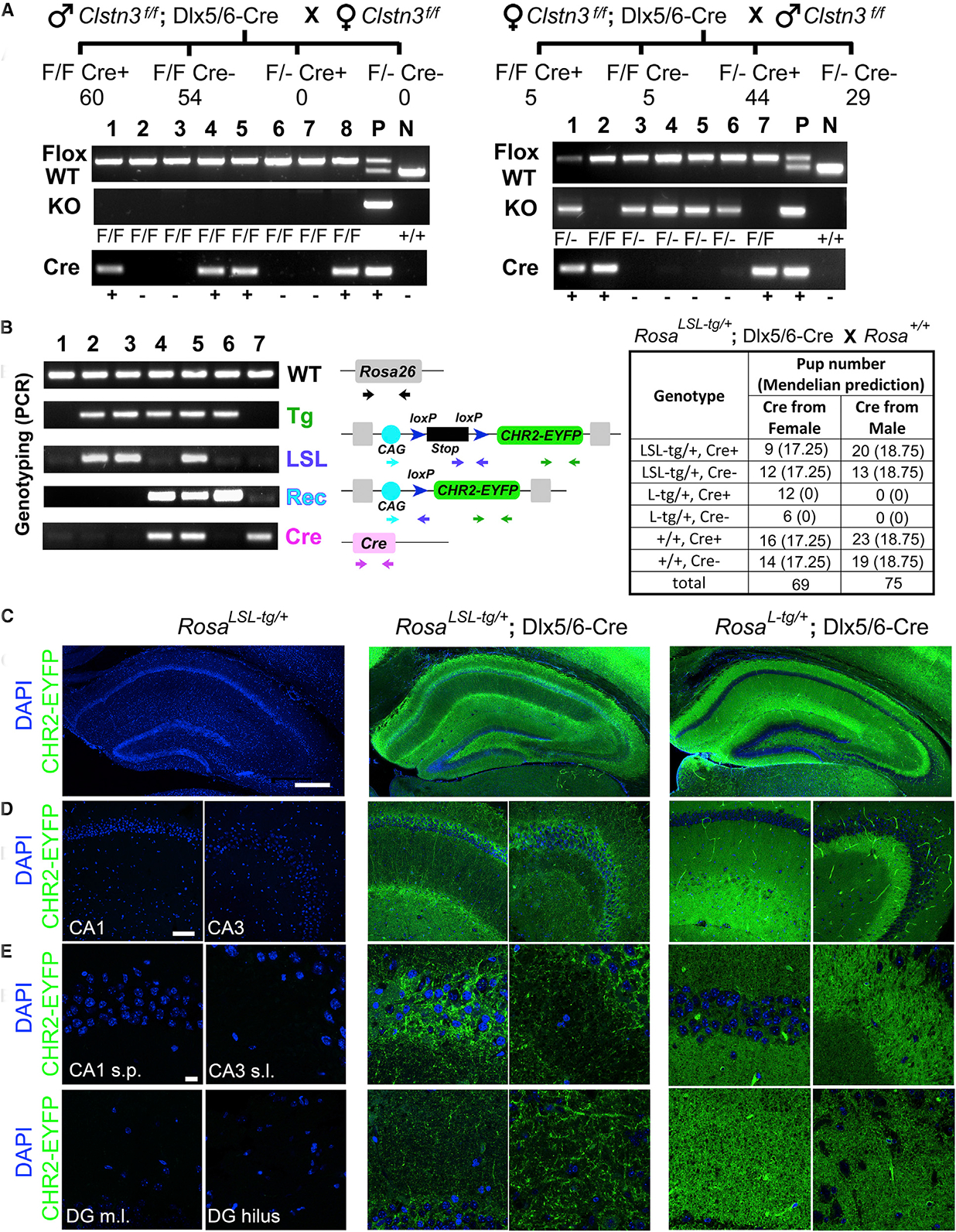
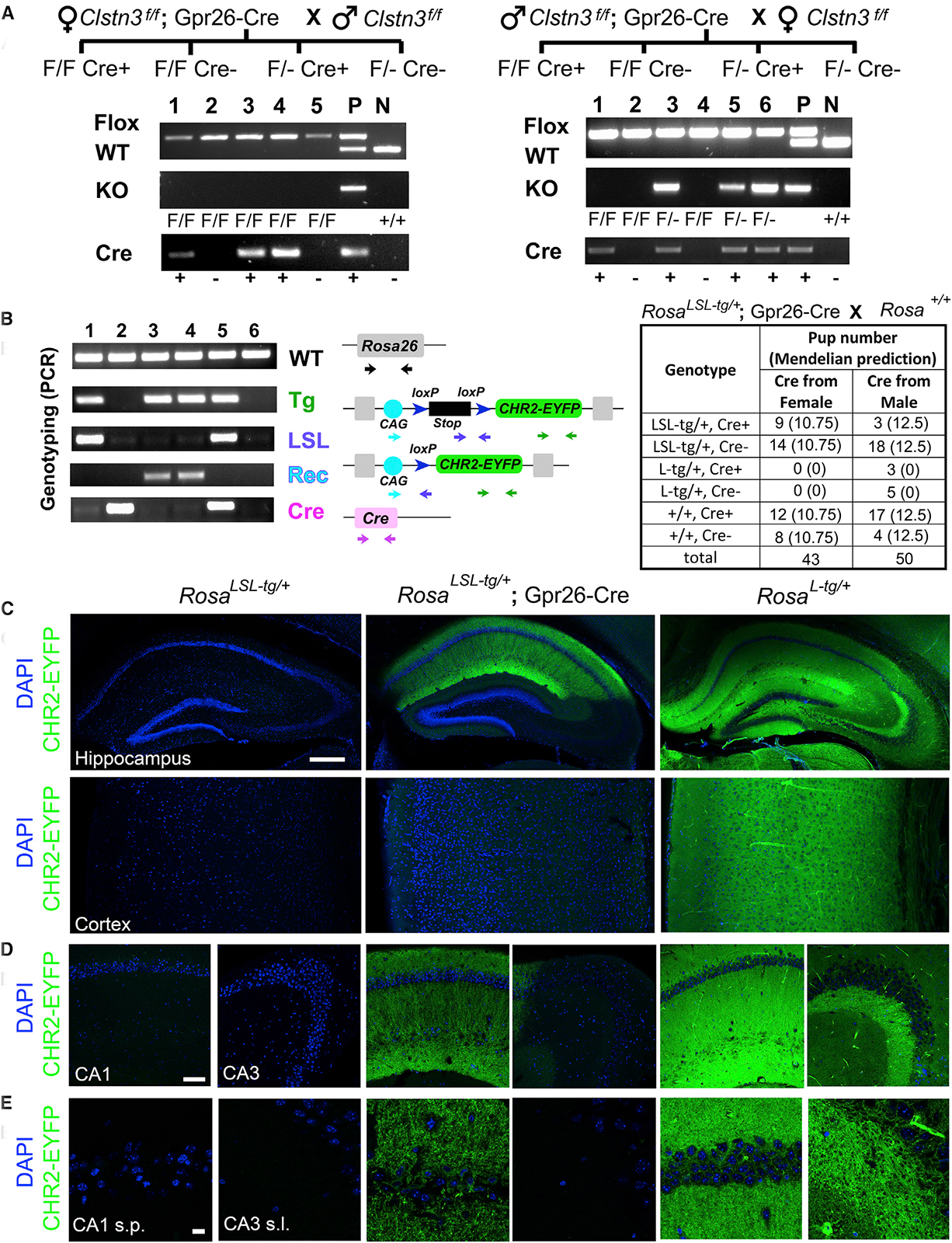
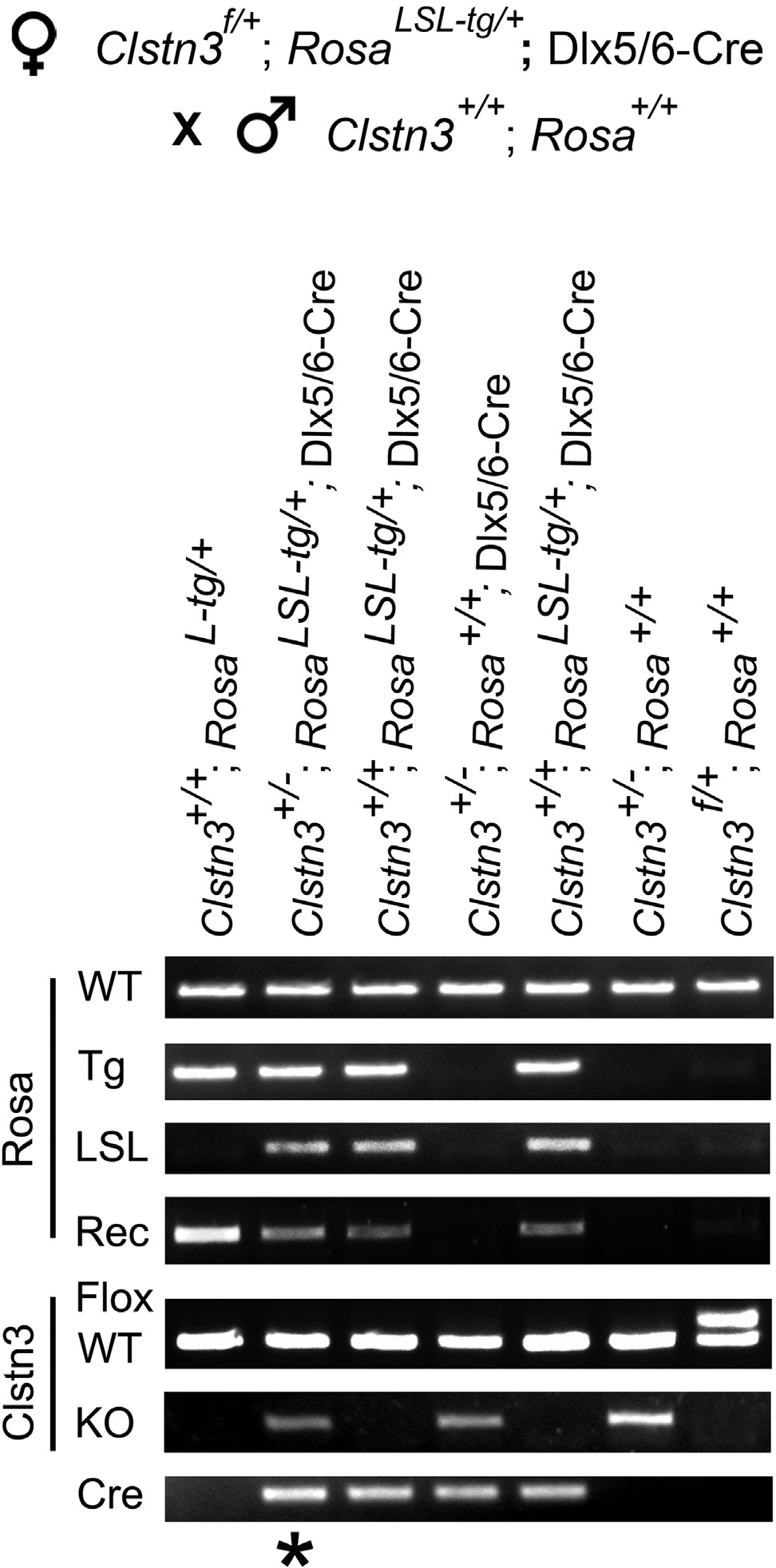
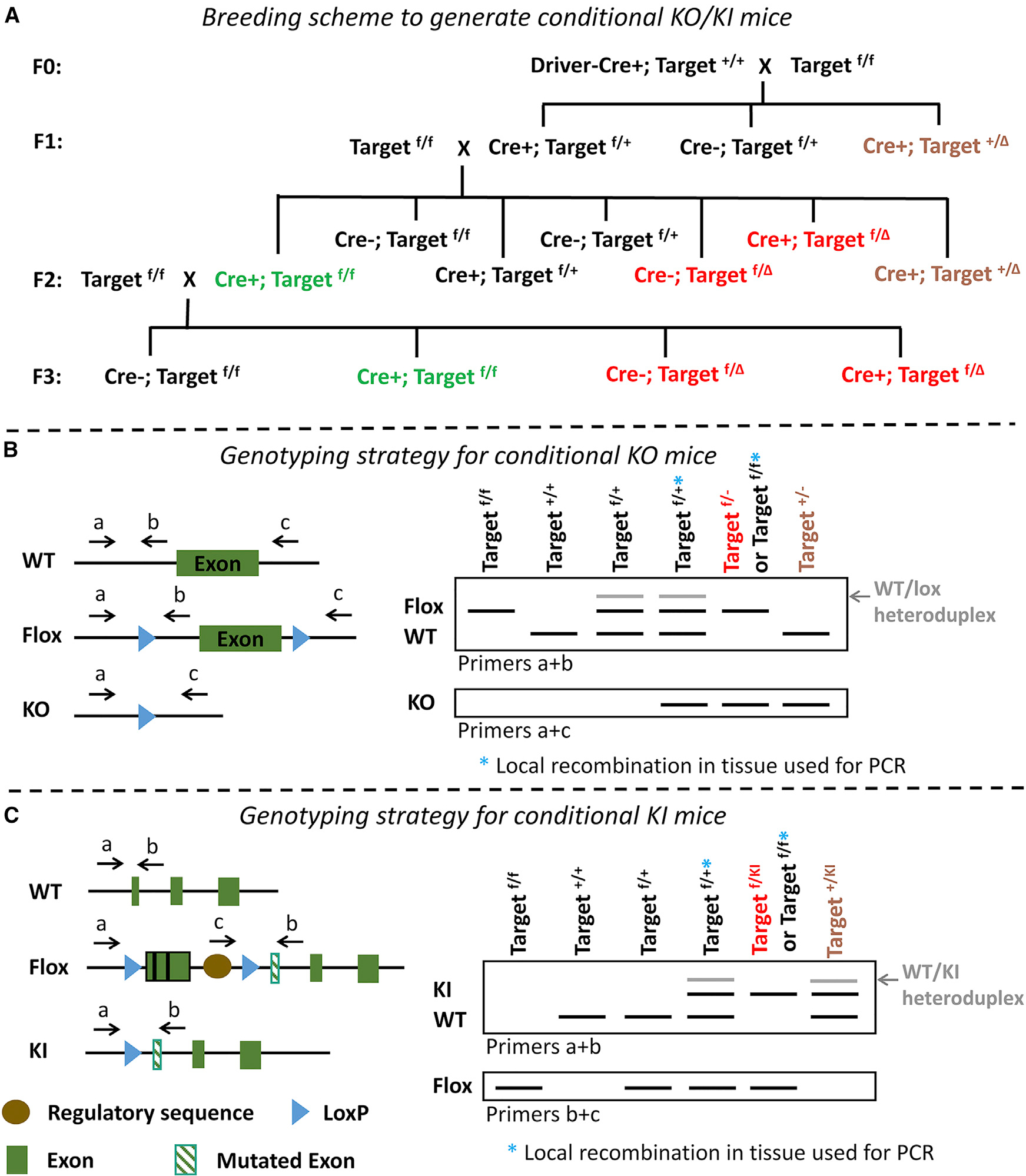
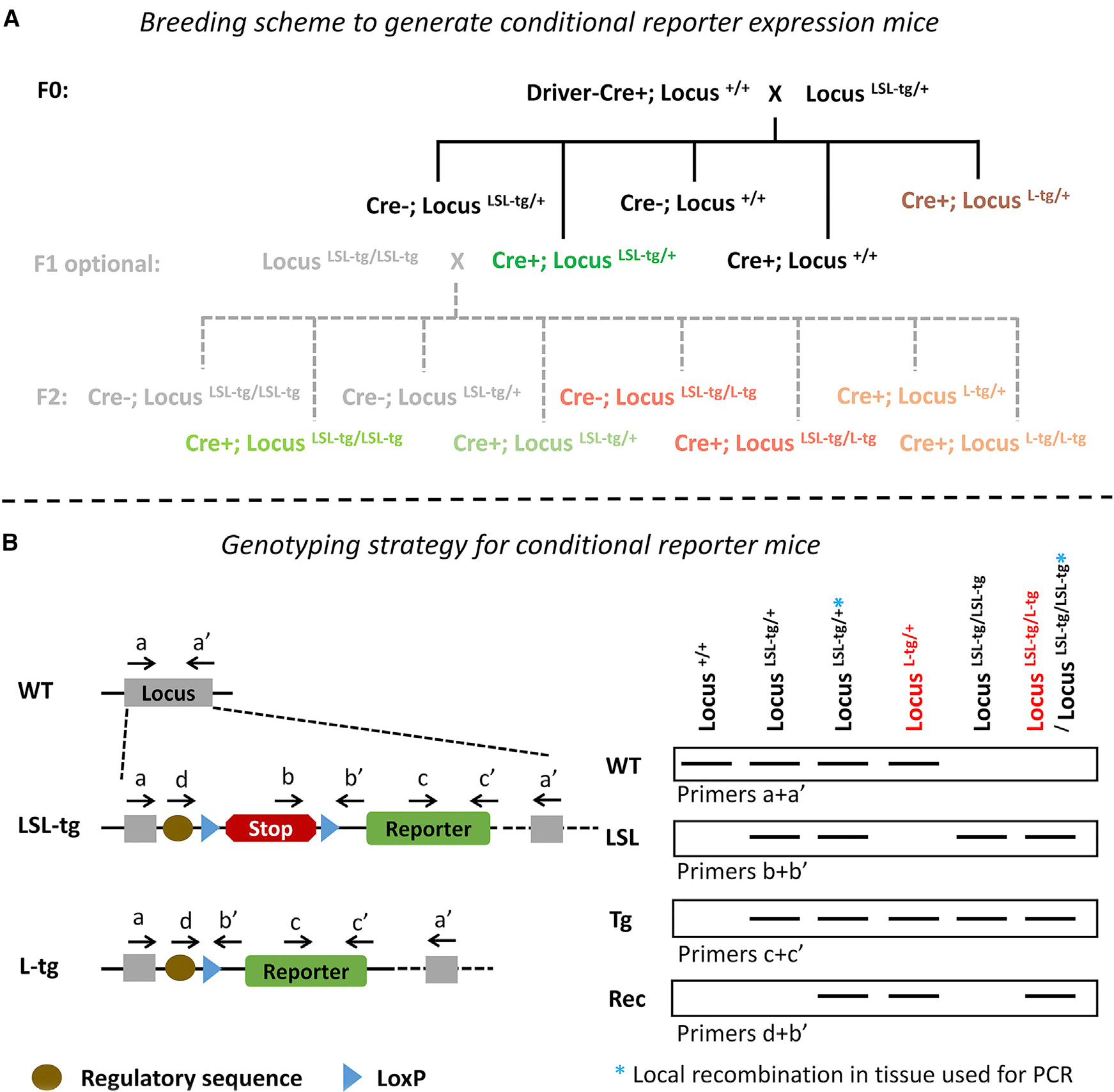
The Cre-loxP system is invaluable for spatial and temporal control of gene knockout, knockin, and reporter expression in the mouse nervous system. However, we report varying probabilities of unexpected germline recombination in distinct Cre driver lines designed for nervous system-specific recombination. Selective maternal or paternal germline recombination is showcased with sample Cre lines. Collated data reveal germline recombination in over half of 64 commonly used Cre driver lines, in most cases with a parental sex bias related to Cre expression in sperm or oocytes. Slight differences among Cre driver lines utilizing common transcriptional control elements affect germline recombination rates. Specific target loci demonstrated differential recombination; thus, reporters are not reliable proxies for another locus of interest. Similar principles apply to other recombinase systems and other genetically targeted organisms. We hereby draw attention to the prevalence of germline recombination and provide guidelines to inform future research for the neuroscience and broader molecular genetics communities.
Keywords: Cre-lox; conditional gene targeting; conditional knockin; conditional knockout; conditional reporter; germline recombination; molecular genetics; mosaic recombination; parental sex bias; site-specific recombinase.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Akashi K, Kakizaki T, Kamiya H, Fukaya M, Yamasaki M, Abe M, Natsume R, Watanabe M, and Sakimura K (2009). NMDA receptor GluN2B (GluR ε 2/NR2B) subunit is crucial for channel function, postsynaptic macromolecular organization, and actin cytoskeleton at hippocampal CA3 synapses. J. Neurosci 29, 10869–10882. - PMC - PubMed
-
- Ambrozkiewicz MC, Schwark M, Kishimoto-Suga M, Borisova E, Hori K, Salazar-Lázaro A, Rusanova A, Altas B, Piepkorn L, Bessa P, et al. (2018). Polarity Acquisition in Cortical Neurons Is Driven by Synergistic Action of Sox9-Regulated Wwp1 and Wwp2 E3 Ubiquitin Ligases and Intronic miR-140. Neuron 100, 1097–1115. - PubMed
-
- Bäckman CM, Malik N, Zhang Y, Shan L, Grinberg A, Hoffer BJ, Westphal H, and Tomac AC (2006). Characterization of a mouse strain expressing Cre recombinase from the 3’ untranslated region of the dopamine transporter locus. Genesis 44, 383–390. - PubMed
-
- Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T, Ferreira M, Tang V, McGovern RA, Kenny CD, et al. (2005). Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell 123, 493–505. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R00 DA041445/DA/NIDA NIH HHS/United States
- P30 EY005722/EY/NEI NIH HHS/United States
- F32 DA038913/DA/NIDA NIH HHS/United States
- R01 EY030611/EY/NEI NIH HHS/United States
- R01 NS057819/NS/NINDS NIH HHS/United States
- R01 NS102237/NS/NINDS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 DA031833/DA/NIDA NIH HHS/United States
- K99 HD092545/HD/NICHD NIH HHS/United States
- R37 MH049428/MH/NIMH NIH HHS/United States
- R01 NS103484/NS/NINDS NIH HHS/United States
- K99 DA041445/DA/NIDA NIH HHS/United States
- R01 EY024694/EY/NEI NIH HHS/United States
- R01 NS083898/NS/NINDS NIH HHS/United States
- R01 MH113349/MH/NIMH NIH HHS/United States
- R01 DA042744/DA/NIDA NIH HHS/United States
- P30 EY012196/EY/NEI NIH HHS/United States
- R01 MH111647/MH/NIMH NIH HHS/United States
- R01 NS050835/NS/NINDS NIH HHS/United States
- R01 NS099099/NS/NINDS NIH HHS/United States
- R01 MH081880/MH/NIMH NIH HHS/United States
- P50 HD103555/HD/NICHD NIH HHS/United States
- R37 NS029169/NS/NINDS NIH HHS/United States
- R01 NS092578/NS/NINDS NIH HHS/United States
- R01 MH049428/MH/NIMH NIH HHS/United States
- F31 NS092419/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
