

Regulation of mesenchymal stem cell differentiation on microstructured titanium surfaces by semaphorin 3A
- PMID: 32028017
- PMCID: PMC7749709
- DOI: 10.1016/j.bone.2020.115260
Regulation of mesenchymal stem cell differentiation on microstructured titanium surfaces by semaphorin 3A
Abstract
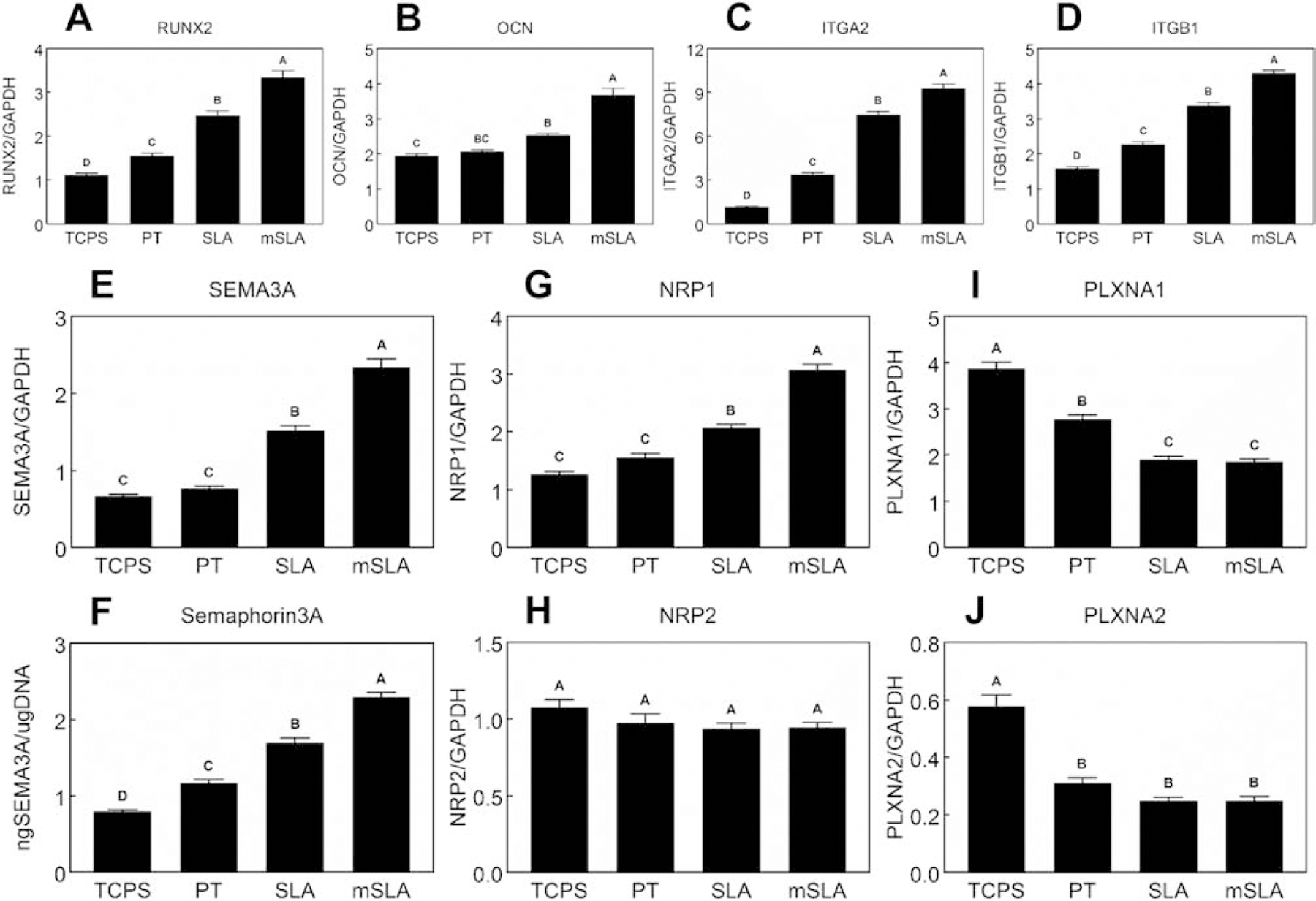
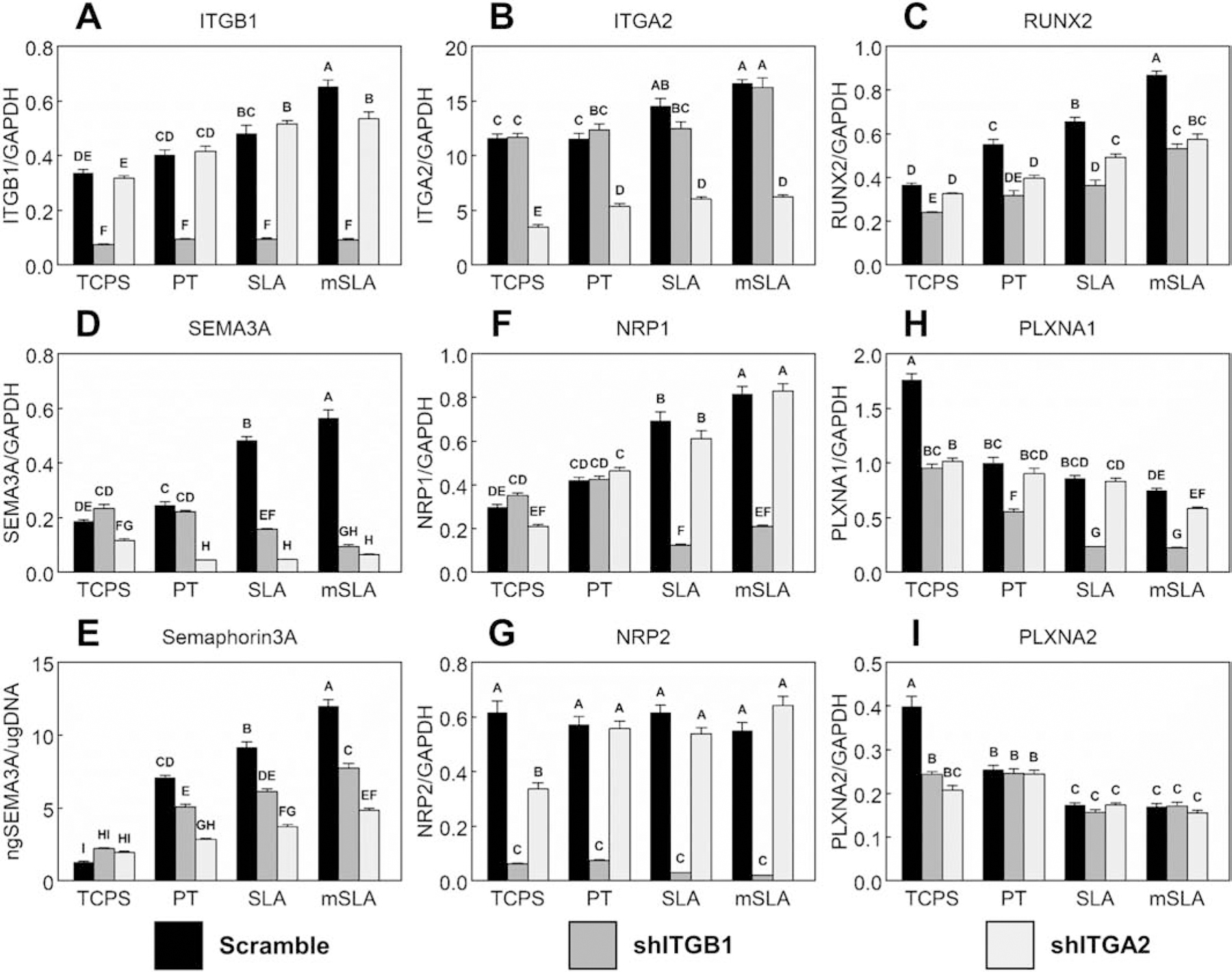
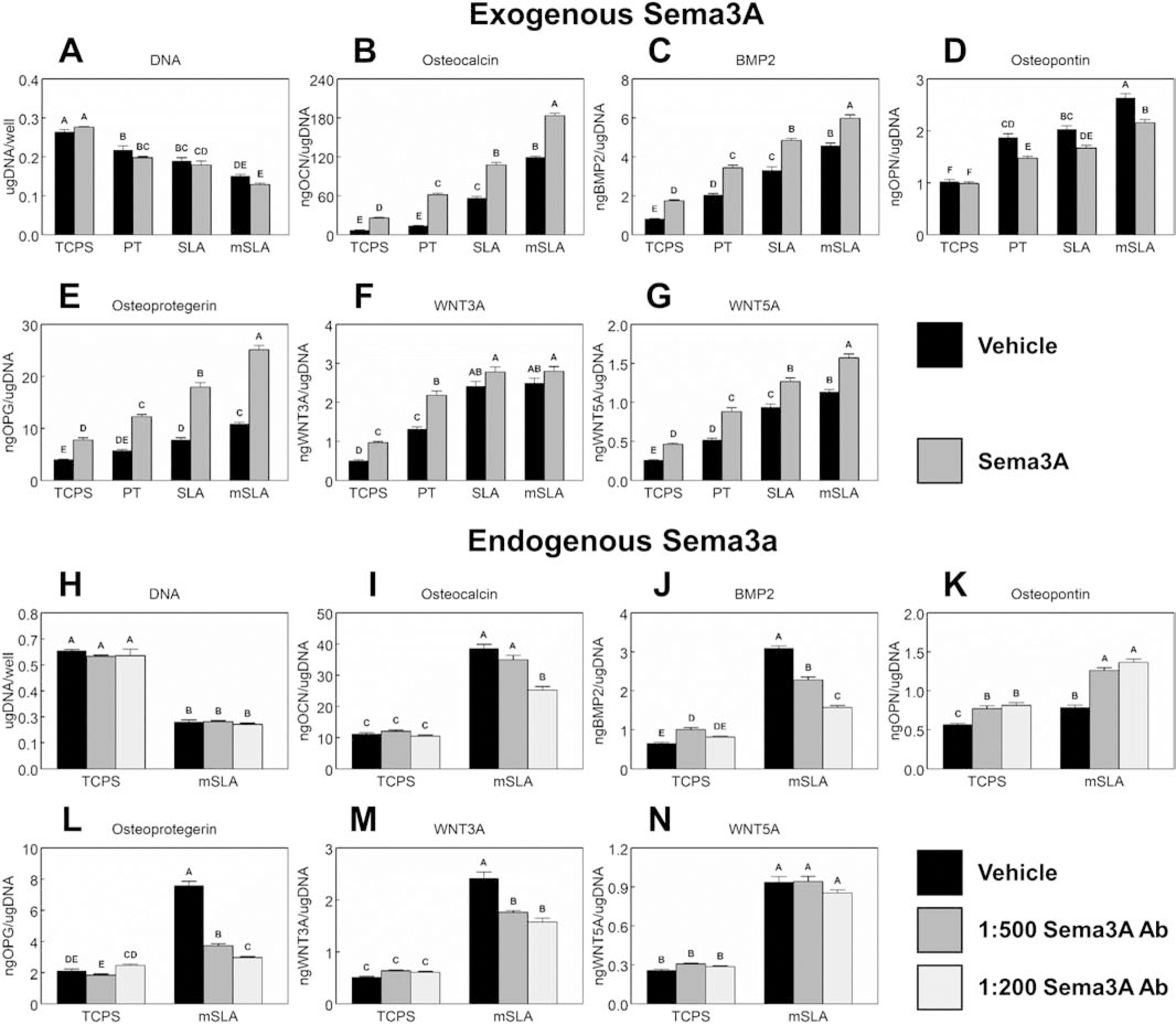
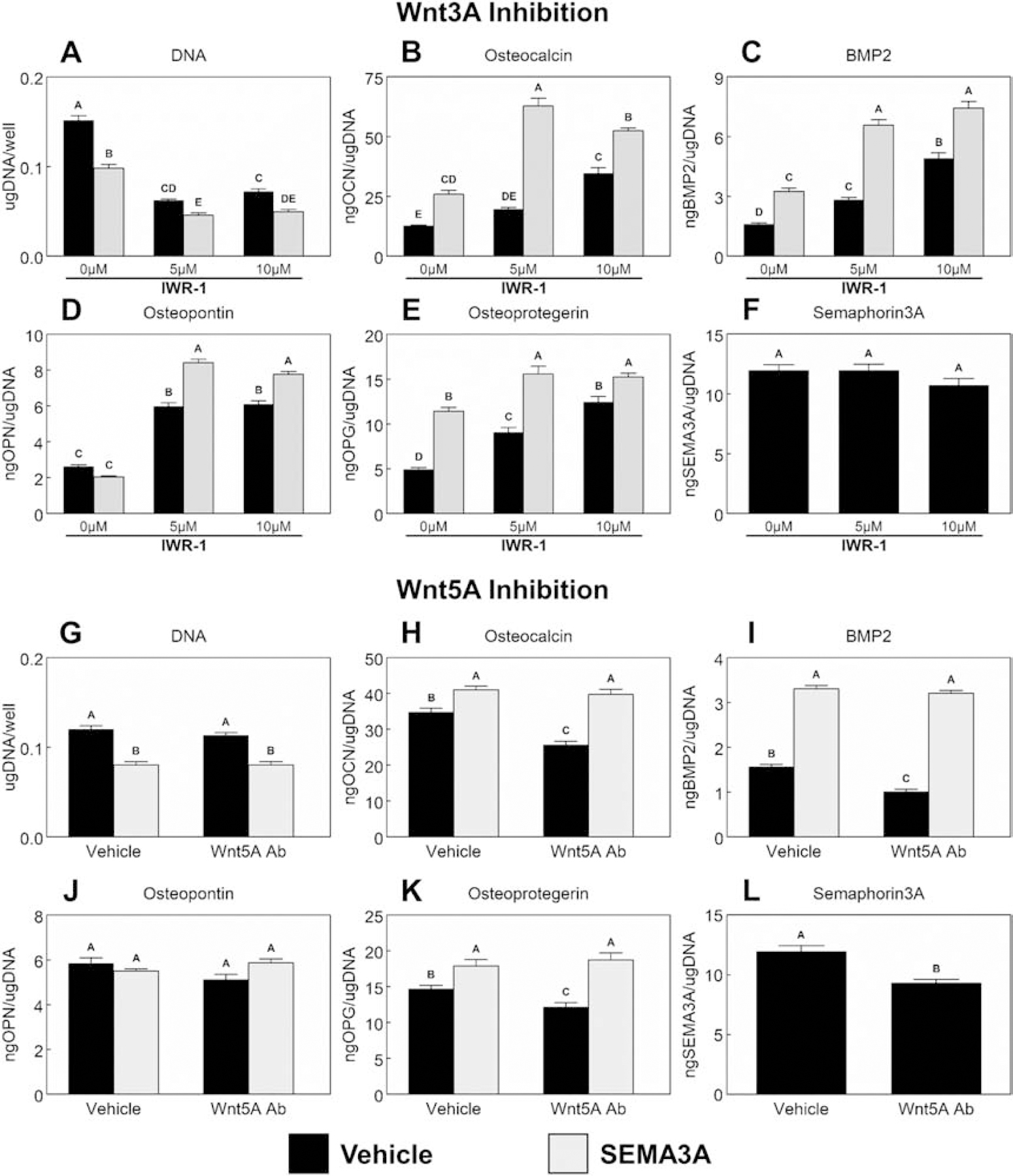
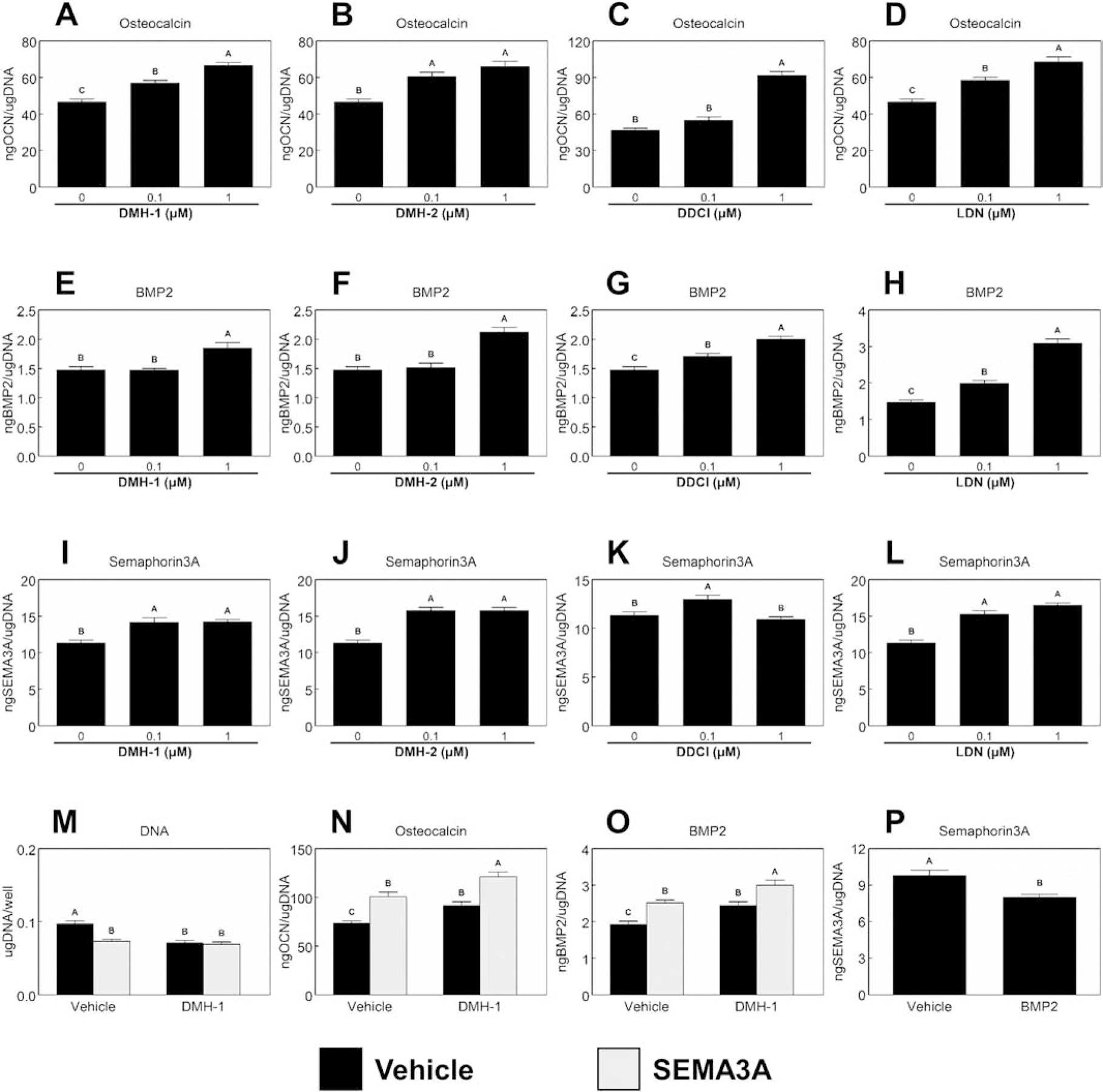
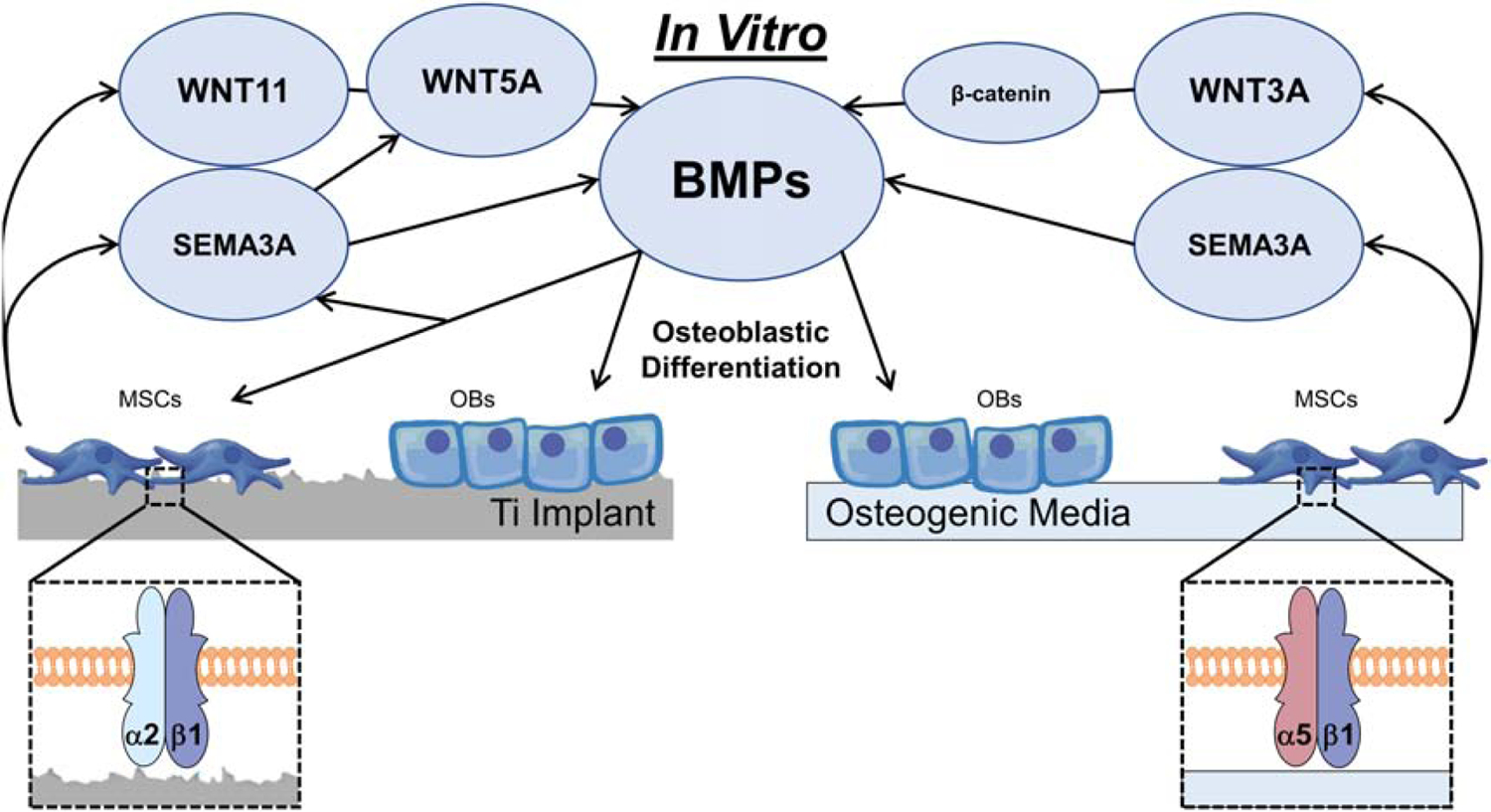
Peri-implant bone formation depends on the ability of mesenchymal stem cells (MSCs) to colonize implant surfaces and differentiate into osteoblasts, but the precise mechanisms controlling this process remain unclear. In vitro, MSCs undergo osteoblastic differentiation on microstructured titanium (Ti) surfaces in the absence of exogenous media supplements and produce factors that promote osteogenesis while regulating osteoclast activity, including semaphorins. The goal of this study was to evaluate the role of semaphorin 3A (Sema3A) on surface-mediated osteoblastic differentiation and determine the hierarchy of this signaling cascade. Human MSCs were cultured on 15 mm grade 2 smooth (pretreatment, PT), hydrophobic-microrough (sand blasted/acid etched, SLA), hydrophilic-microrough Ti (mSLA) (Institut Straumann AG, Basel, Switzerland), or tissue culture polystyrene (TCPS). Expression of SEMA3A family proteins increased after 7 days of culture, and the increased expression in response to microstructured Ti was dependent on recognition of the surface by integrin α2β1. Exogenous Sema3A increased differentiation whereas differentiation was decreased in cells treated with a Sema3A antibody. Furthermore, Sema3A influenced the production of osteoprotegerin and osteopontin suggesting it as an important local regulator of bone remodeling. Inhibition of Wnt3A and Wnt5A revealed that activation of Sema3A occurs downstream of Wnt5A and may facilitate the translocation of β-catenin bypassing the canonical Wnt3A initiating signal associated with osteoblastic differentiation. Furthermore, chemical inhibition of calmodulin (CaM), Ca2+/calmodulin-dependent protein kinase (CaMKII), phospholipase A2 (PLA2), protein kinase C (PKC), and BMP receptors suggest that Sema3A could serve as a feedback mechanism for both Wnt5A and BMP2. Here, we show novel roles for Sema3A family proteins in the surface-dependent modulation of MSCs as well as important interactions with pathways known to be associated with osteoblastic differentiation. Moreover, their effects on bone remodeling markers have significant implications for peri-implant bone remodeling and downstream modulation of osteoclastic activity. These results suggest that Sema3A aids in peri-implant bone formation through regulation on multiple stages of osseointegration, making it a potential target to promote osseointegration in patients with compromised bone remodeling.
Keywords: Implant; Mesenchymal stem cell; Osteoblast; Semaphorin3A; Surface topography; Titanium.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
