

PARPs and ADP-ribosylation in RNA biology: from RNA expression and processing to protein translation and proteostasis
- PMID: 32029452
- PMCID: PMC7050490
- DOI: 10.1101/gad.334433.119
PARPs and ADP-ribosylation in RNA biology: from RNA expression and processing to protein translation and proteostasis
Abstract
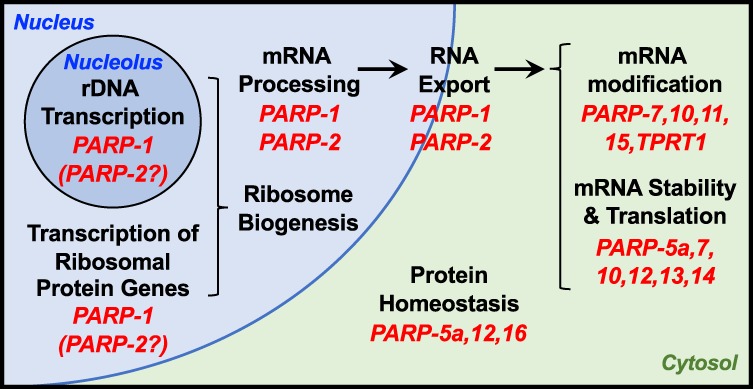
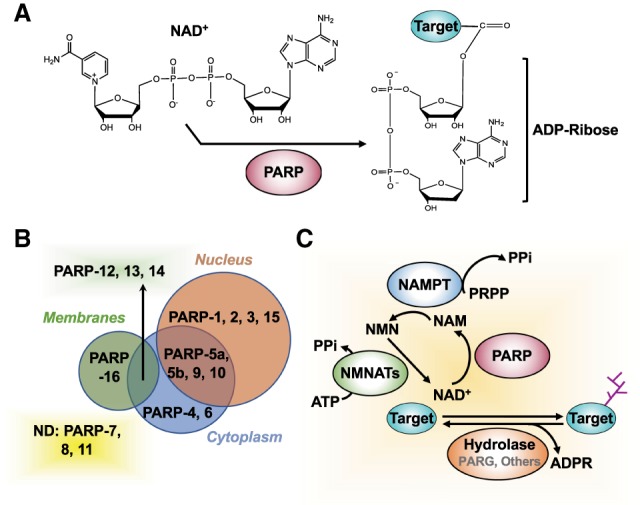
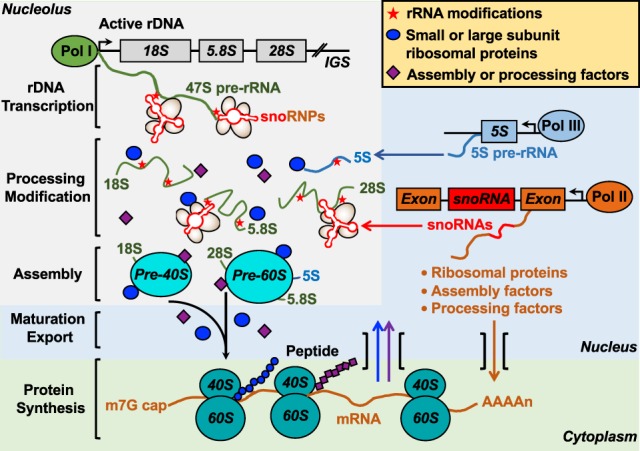
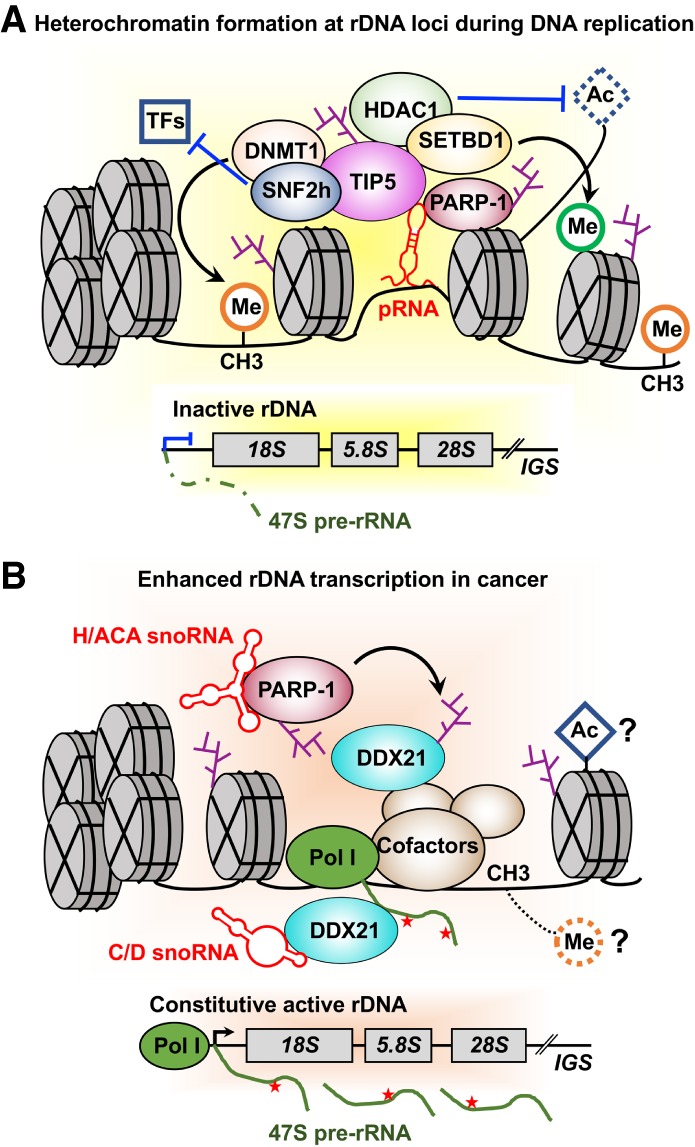
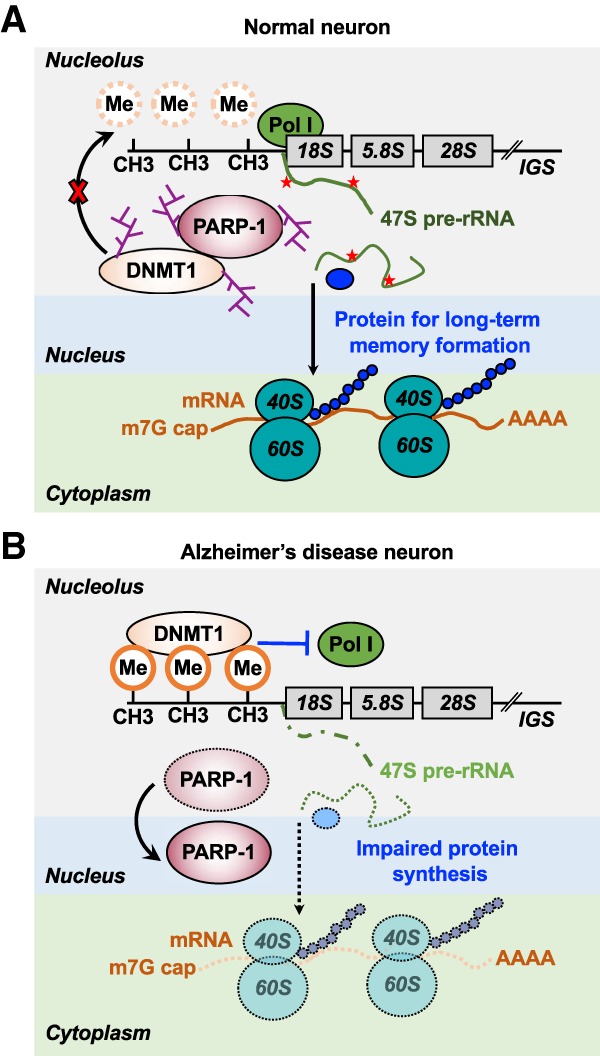
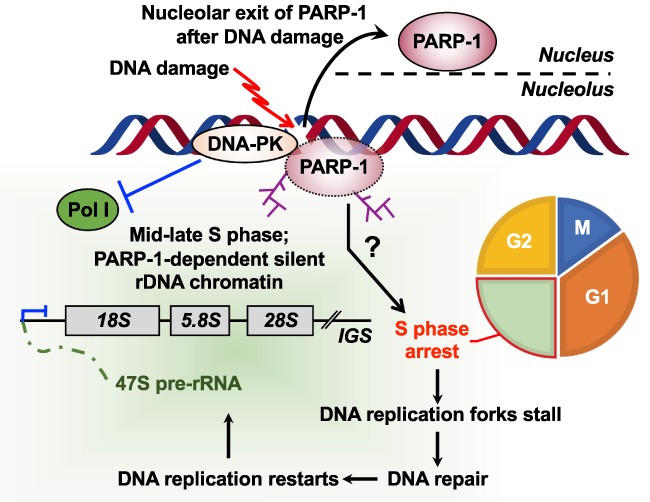
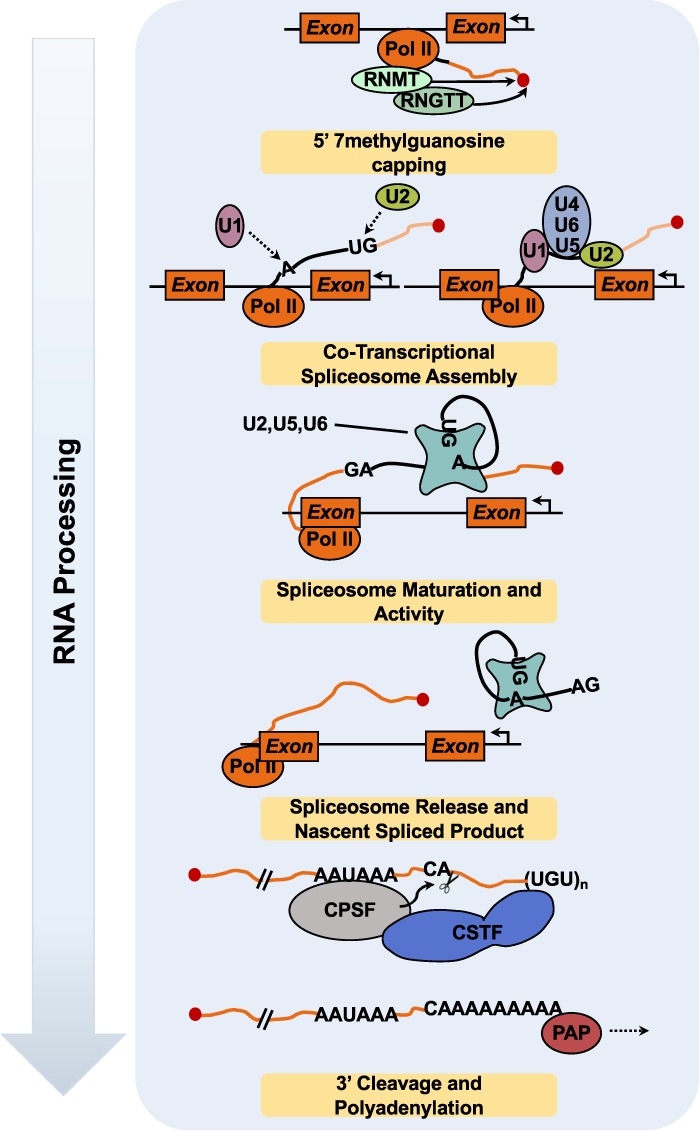
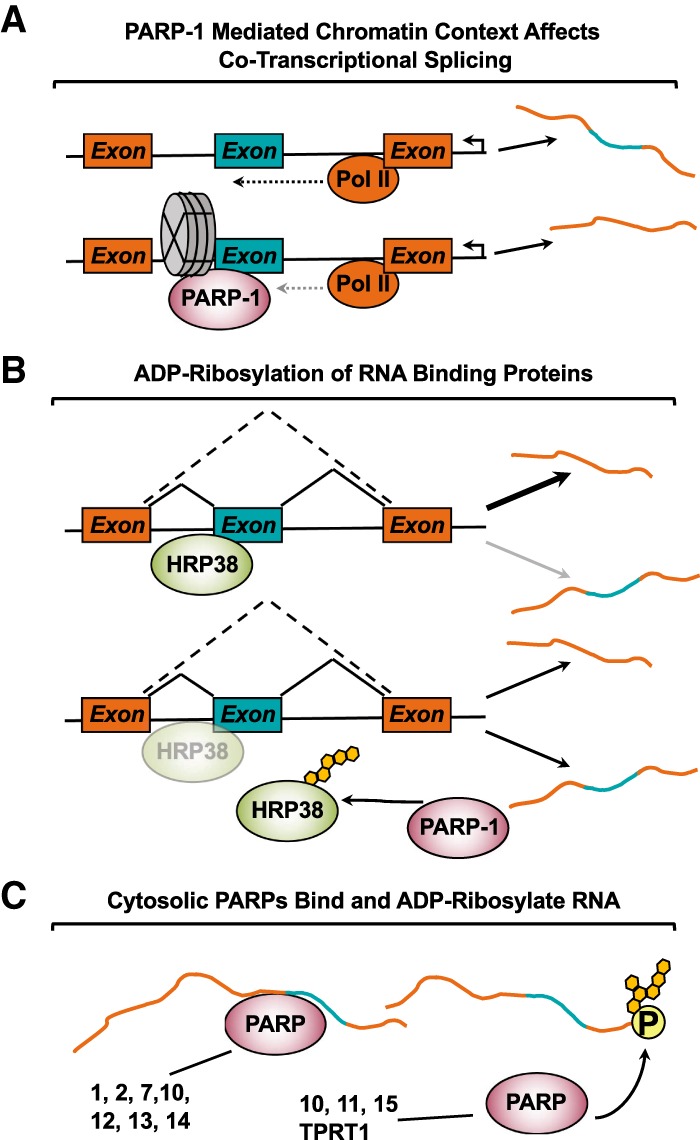
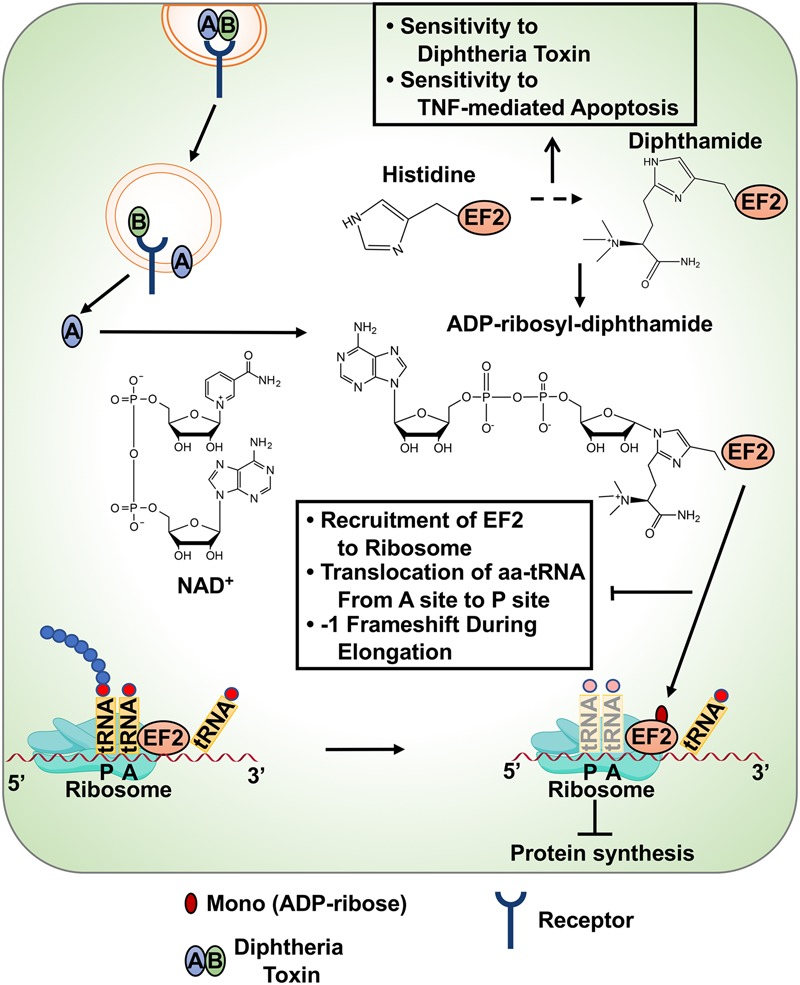
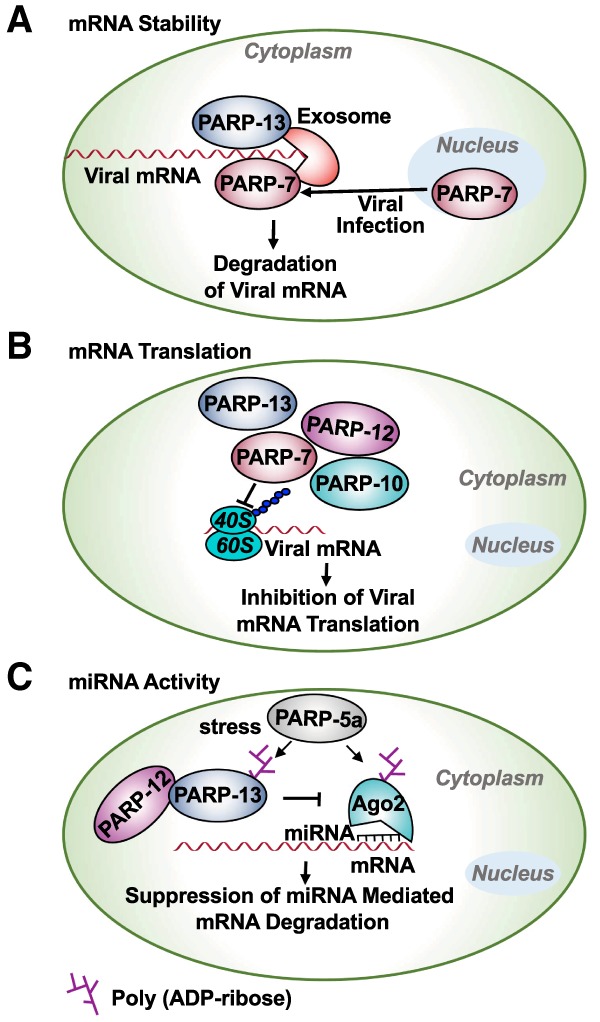
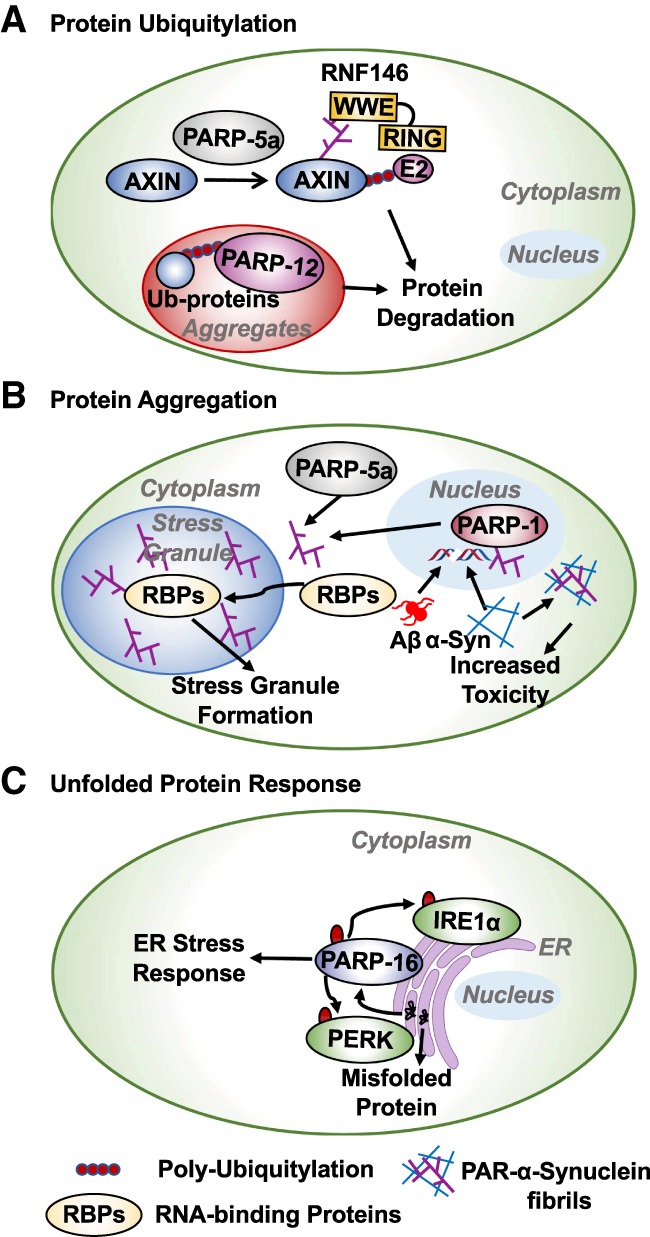
ADP-ribosylation (ADPRylation) is a posttranslational modification of proteins discovered nearly six decades ago, but many important questions remain regarding its molecular functions and biological roles, as well as the activity of the ADP-ribose (ADPR) transferase enzymes (PARP family members) that catalyze it. Growing evidence indicates that PARP-mediated ADPRylation events are key regulators of the protein biosynthetic pathway, leading from rDNA transcription and ribosome biogenesis to mRNA synthesis, processing, and translation. In this review we describe the role of PARP proteins and ADPRylation in all facets of this pathway. PARP-1 and its enzymatic activity are key regulators of rDNA transcription, which is a critical step in ribosome biogenesis. An emerging role of PARPs in alternative splicing of mRNAs, as well as direct ADPRylation of mRNAs, highlight the role of PARP members in RNA processing. Furthermore, PARP activity, stimulated by cellular stresses, such as viral infections and ER stress, leads to the regulation of mRNA stability and protein synthesis through posttranscriptional mechanisms. Dysregulation of PARP activity in these processes can promote disease states. Collectively, these results highlight the importance of PARP family members and ADPRylation in gene regulation, mRNA processing, and protein abundance. Future studies in these areas will yield new insights into the fundamental mechanisms and a broader utility for PARP-targeted therapeutic agents.
Keywords: ADP-ribosylation (ADPRylation); DNA damage; PARP inhibitors (PARPi); RNA stability; mRNA processing; mRNA splicing; mRNA translation; mono(ADP-ribose) (MAR); poly(ADP-ribose) (PAR); poly(ADP-ribose) polymerase (PARP); rRNA synthesis; ribosome biogenesis; stress responses.
© 2020 Kim et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Abraham R, Hauer D, McPherson RL, Utt A, Kirby IT, Cohen MS, Merits A, Leung AKL, Griffin DE. 2018. ADP-ribosyl-binding and hydrolase activities of the alphavirus nsP3 macrodomain are critical for initiation of virus replication. Proc Natl Acad Sci 115: E10457–E10466. 10.1073/pnas.1812130115 - DOI - PMC - PubMed
-
- Ahmed S, Bott D, Gomez A, Tamblyn L, Rasheed A, Cho T, MacPherson L, Sugamori KS, Yang Y, Grant DM, et al. 2015. Loss of the mono-ADP-ribosyltransferase, Tiparp, increases sensitivity to dioxin-induced steatohepatitis and lethality. J Biol Chem 290: 16824–16840. 10.1074/jbc.M115.660100 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous