

The impact of PARPs and ADP-ribosylation on inflammation and host-pathogen interactions
- PMID: 32029454
- PMCID: PMC7050484
- DOI: 10.1101/gad.334425.119
The impact of PARPs and ADP-ribosylation on inflammation and host-pathogen interactions
Abstract
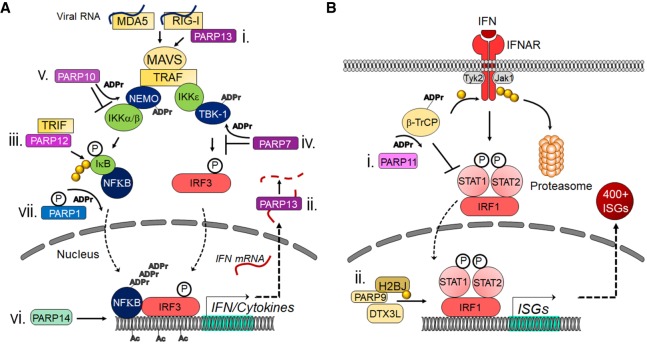
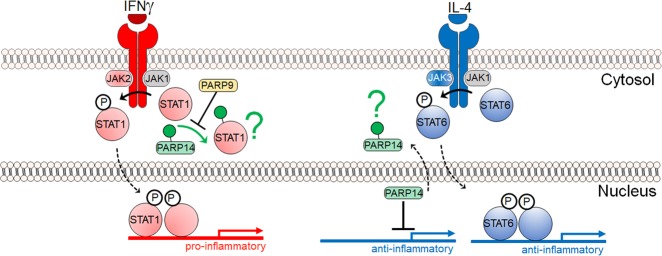
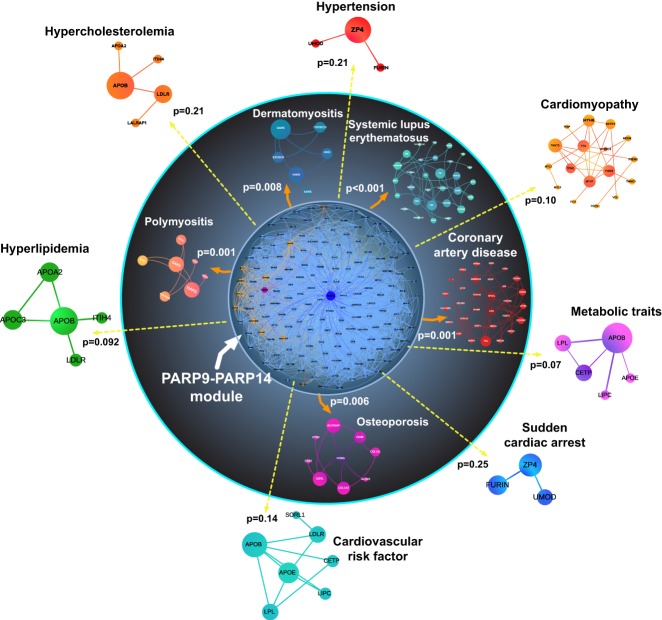
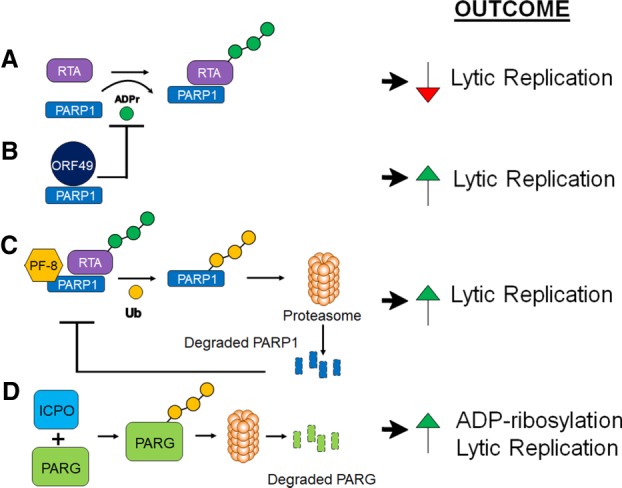
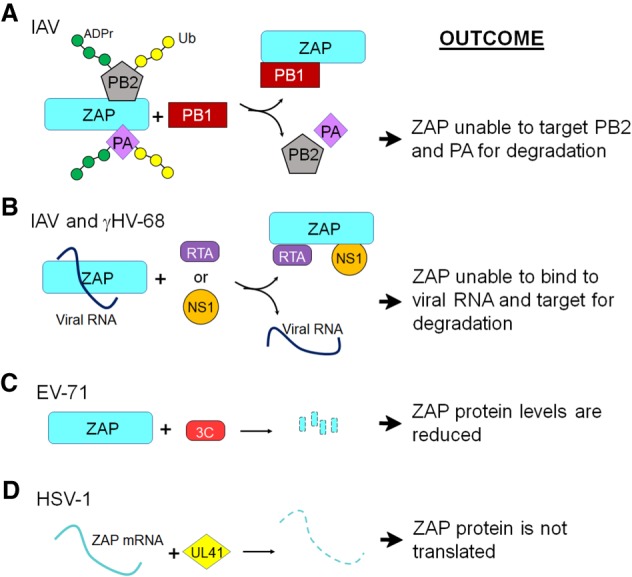
Poly-adenosine diphosphate-ribose polymerases (PARPs) promote ADP-ribosylation, a highly conserved, fundamental posttranslational modification (PTM). PARP catalytic domains transfer the ADP-ribose moiety from NAD+ to amino acid residues of target proteins, leading to mono- or poly-ADP-ribosylation (MARylation or PARylation). This PTM regulates various key biological and pathological processes. In this review, we focus on the roles of the PARP family members in inflammation and host-pathogen interactions. Here we give an overview the current understanding of the mechanisms by which PARPs promote or suppress proinflammatory activation of macrophages, and various roles PARPs play in virus infections. We also demonstrate how innovative technologies, such as proteomics and systems biology, help to advance this research field and describe unanswered questions.
Keywords: ADP-ribosylation; PARP; atherosclerosis; host–pathogen interactions; immunity; inflammation; macrophage; vascular disease.
© 2020 Fehr et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Abraham R, Hauer D, McPherson RL, Utt A, Kirby IT, Cohen MS, Merits A, Leung AKL, Griffin DE. 2018. ADP-ribosyl-binding and hydrolase activities of the alphavirus nsP3 macrodomain are critical for initiation of virus replication. Proc Natl Acad Sci 115: E10457–E10466. 10.1073/pnas.1812130115 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases