

Defective HIV-1 proviruses produce viral proteins
- PMID: 32029589
- PMCID: PMC7035625
- DOI: 10.1073/pnas.1917876117
Defective HIV-1 proviruses produce viral proteins
Abstract
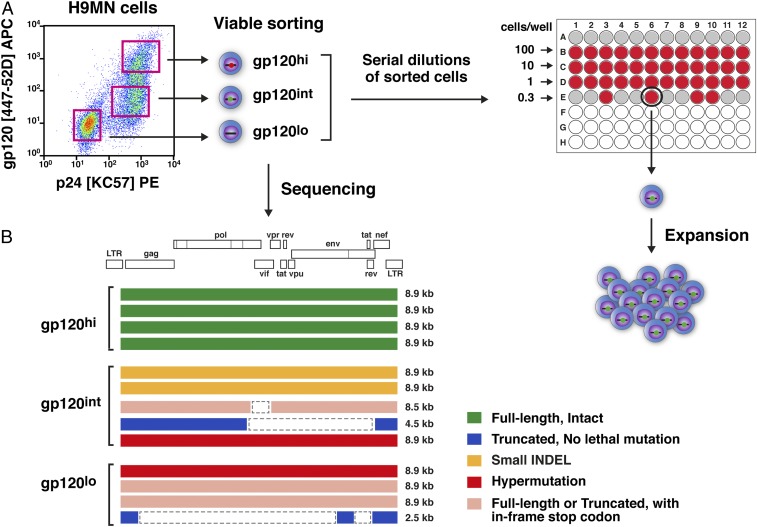
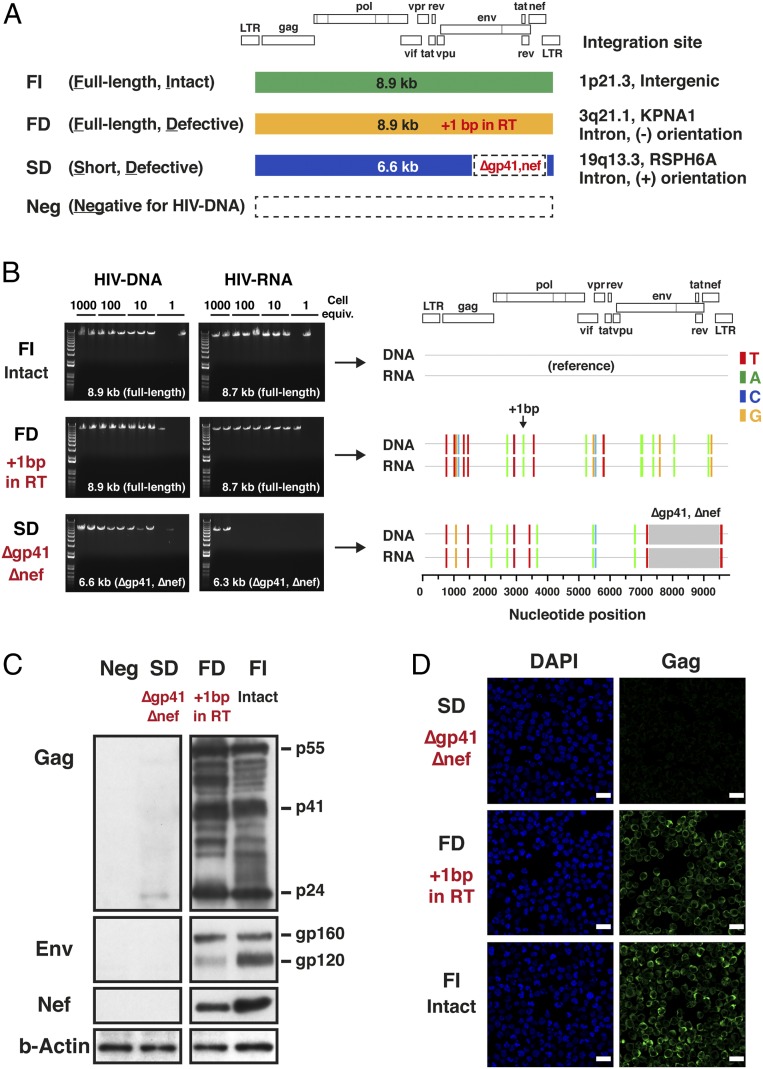
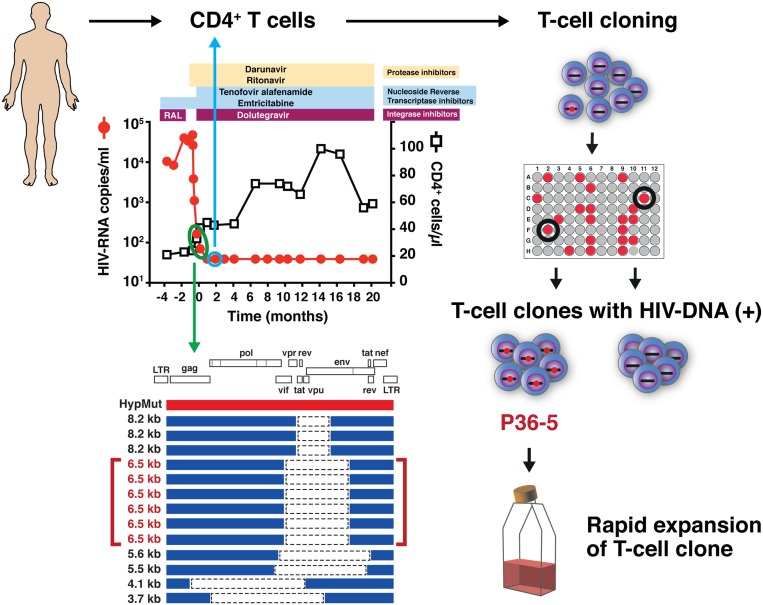
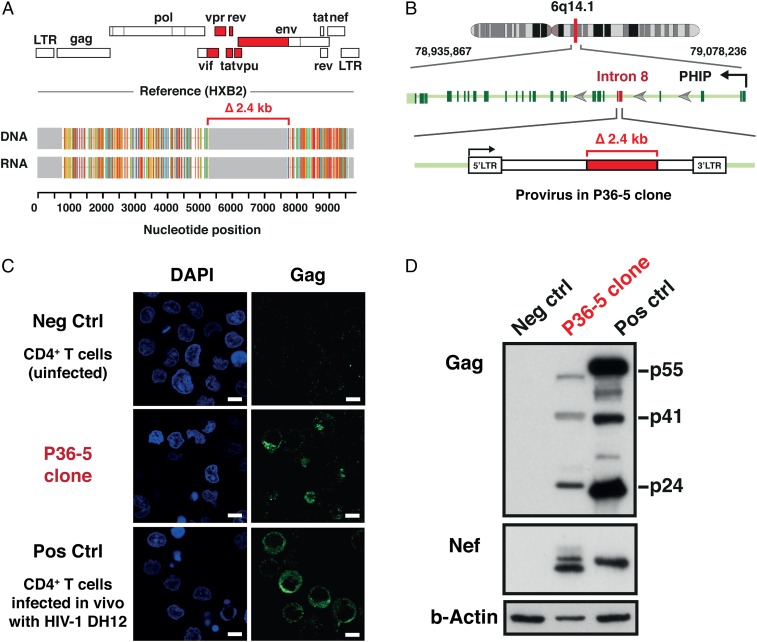
HIV-1 proviruses persist in the CD4+ T cells of HIV-infected individuals despite years of combination antiretroviral therapy (cART) with suppression of HIV-1 RNA levels <40 copies/mL. Greater than 95% of these proviruses detected in circulating peripheral blood mononuclear cells (PBMCs) are referred to as "defective" by virtue of having large internal deletions and lethal genetic mutations. As these defective proviruses are unable to encode intact and replication-competent viruses, they have long been thought of as biologically irrelevant "graveyard" of viruses with little significance to HIV-1 pathogenesis. Contrary to this notion, we have recently demonstrated that these defective proviruses are not silent, are capable of transcribing novel unspliced forms of HIV-RNA transcripts with competent open reading frames (ORFs), and can be found in the peripheral blood CD4+ T cells of patients at all stages of HIV-1 infection. In the present study, by an approach of combining serial dilutions of CD4+ T cells and T cell-cloning technologies, we are able to demonstrate that defective proviruses that persist in HIV-infected individuals during suppressive cART are translationally competent and produce the HIV-1 Gag and Nef proteins. The HIV-RNA transcripts expressed from these defective proviruses may trigger an element of innate immunity. Likewise, the viral proteins coded in the defective proviruses may form extracellular virus-like particles and may trigger immune responses. The persistent production of HIV-1 proteins in the absence of viral replication helps explain persistent immune activation despite HIV-1 levels below detection, and also presents new challenges to HIV-1 eradication.
Keywords: HIV; immune activation; provirus.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Imai H., et al. , A defective proviral DNA with a 2.6-kb deletion of human immunodeficiency virus type 1 (HIV-1) in a persistently HIV-1 infected cell clone. Virus Genes 5, 81–88 (1991). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
