Skp, Cullin, F-box (SCF)-Met30 and SCF-Cdc4-Mediated Proteolysis of CENP-A Prevents Mislocalization of CENP-A for Chromosomal Stability in Budding Yeast
- PMID: 32032354
- PMCID: PMC7032732
- DOI: 10.1371/journal.pgen.1008597
Skp, Cullin, F-box (SCF)-Met30 and SCF-Cdc4-Mediated Proteolysis of CENP-A Prevents Mislocalization of CENP-A for Chromosomal Stability in Budding Yeast
Abstract
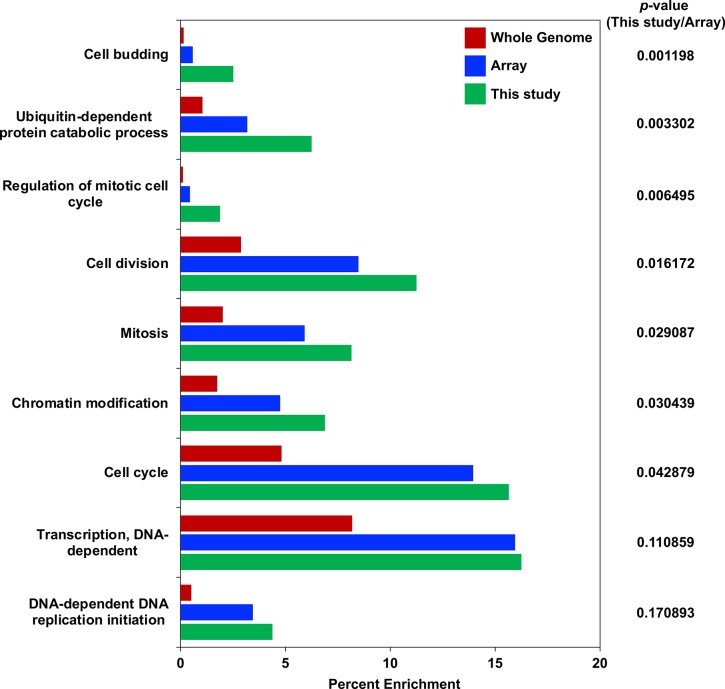
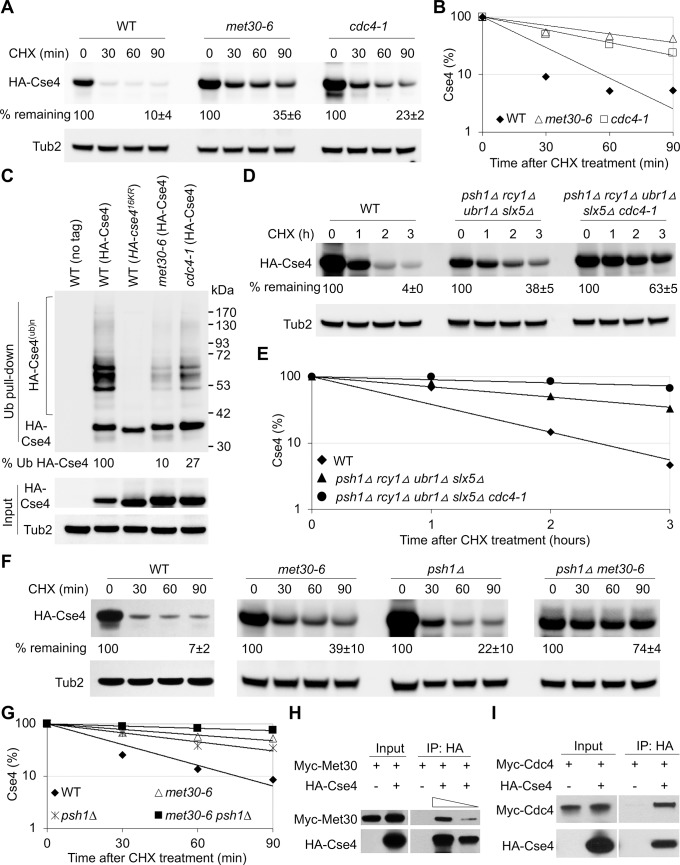
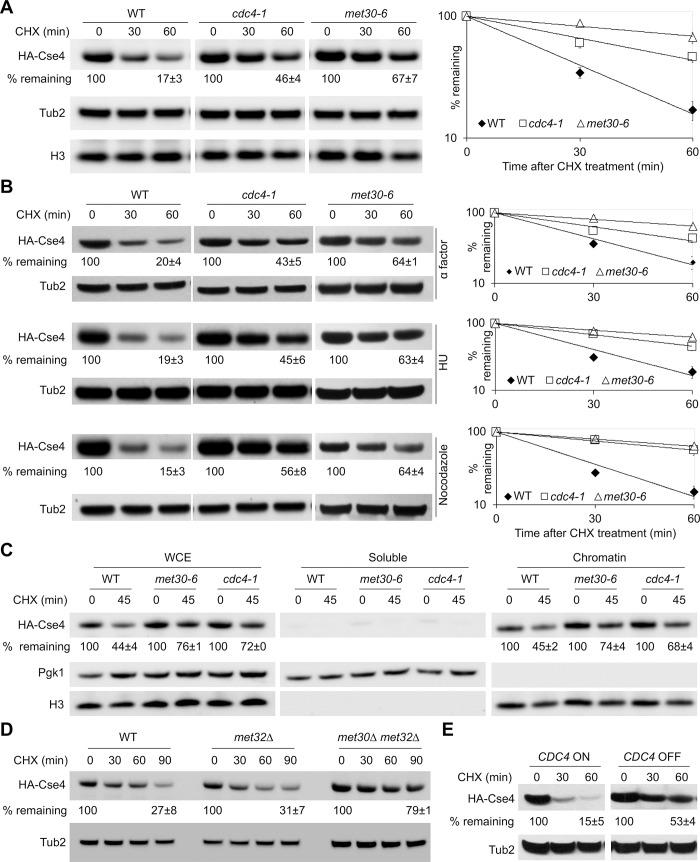
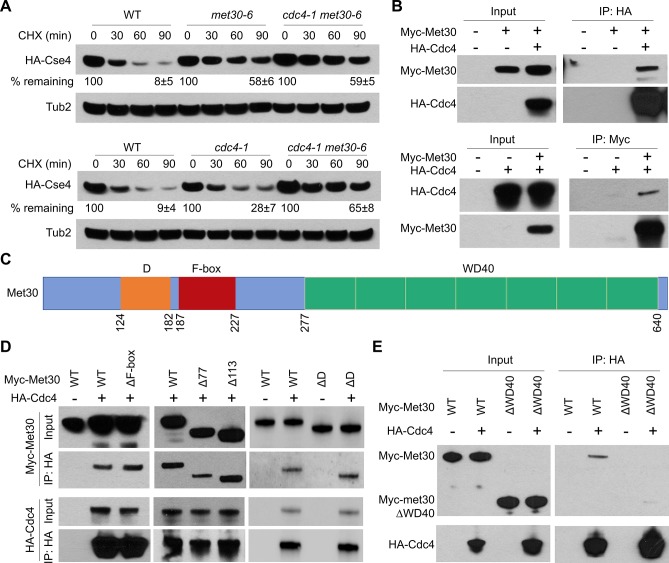
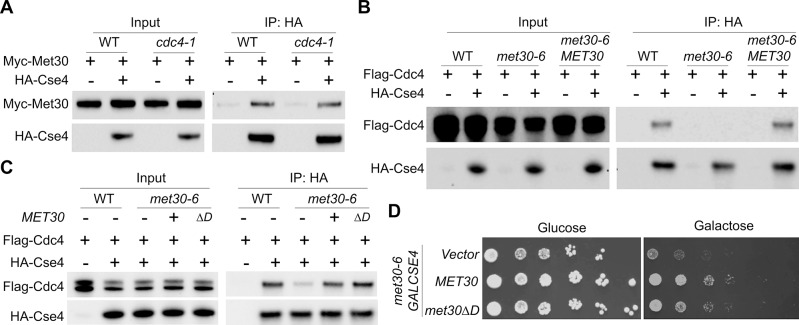
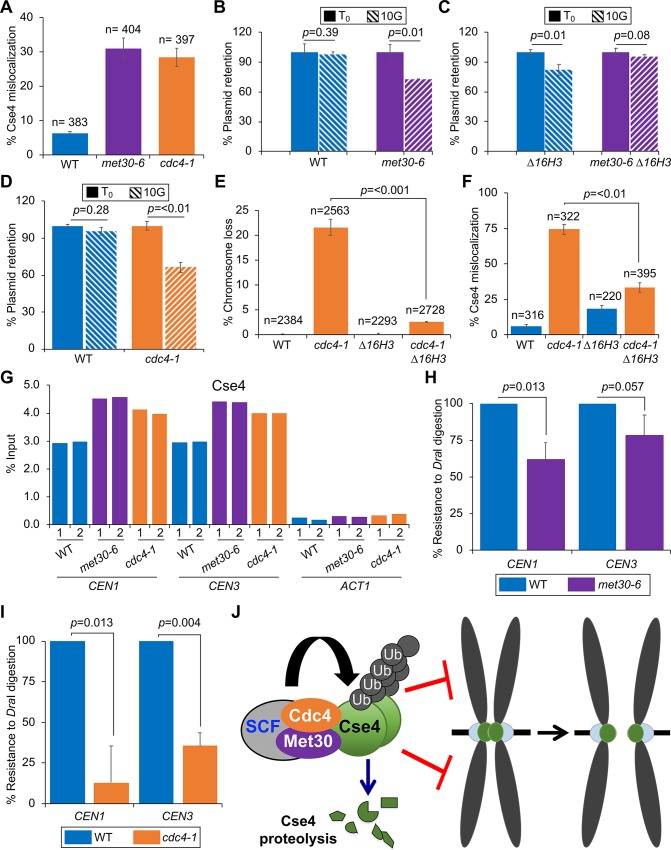
Restricting the localization of the histone H3 variant CENP-A (Cse4 in yeast, CID in flies) to centromeres is essential for faithful chromosome segregation. Mislocalization of CENP-A leads to chromosomal instability (CIN) in yeast, fly and human cells. Overexpression and mislocalization of CENP-A has been observed in many cancers and this correlates with increased invasiveness and poor prognosis. Yet genes that regulate CENP-A levels and localization under physiological conditions have not been defined. In this study we used a genome-wide genetic screen to identify essential genes required for Cse4 homeostasis to prevent its mislocalization for chromosomal stability. We show that two Skp, Cullin, F-box (SCF) ubiquitin ligases with the evolutionarily conserved F-box proteins Met30 and Cdc4 interact and cooperatively regulate proteolysis of endogenous Cse4 and prevent its mislocalization for faithful chromosome segregation under physiological conditions. The interaction of Met30 with Cdc4 is independent of the D domain, which is essential for their homodimerization and ubiquitination of other substrates. The requirement for both Cdc4 and Met30 for ubiquitination is specifc for Cse4; and a common substrate for Cdc4 and Met30 has not previously been described. Met30 is necessary for the interaction between Cdc4 and Cse4, and defects in this interaction lead to stabilization and mislocalization of Cse4, which in turn contributes to CIN. We provide the first direct link between Cse4 mislocalization to defects in kinetochore structure and show that SCF-mediated proteolysis of Cse4 is a major mechanism that prevents stable maintenance of Cse4 at non-centromeric regions, thus ensuring faithful chromosome segregation. In summary, we have identified essential pathways that regulate cellular levels of endogenous Cse4 and shown that proteolysis of Cse4 by SCF-Met30/Cdc4 prevents mislocalization and CIN in unperturbed cells.
Conflict of interest statement
NO authors have competing interests.
Figures