RNA decay during gammaherpesvirus infection reduces RNA polymerase II occupancy of host promoters but spares viral promoters
- PMID: 32032393
- PMCID: PMC7032723
- DOI: 10.1371/journal.ppat.1008269
RNA decay during gammaherpesvirus infection reduces RNA polymerase II occupancy of host promoters but spares viral promoters
Abstract
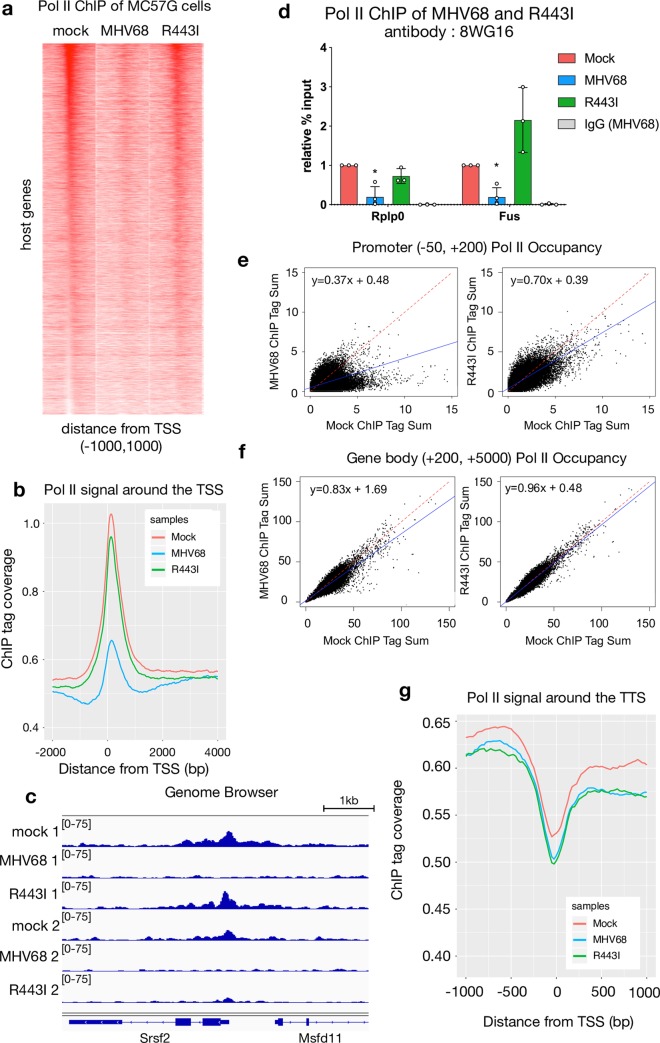
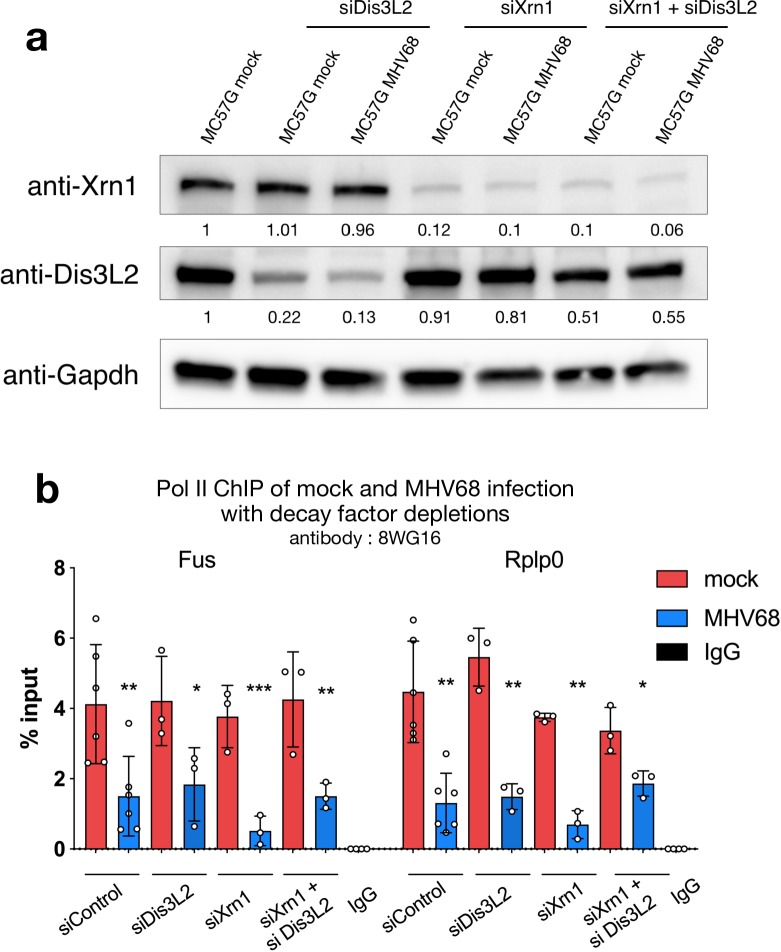
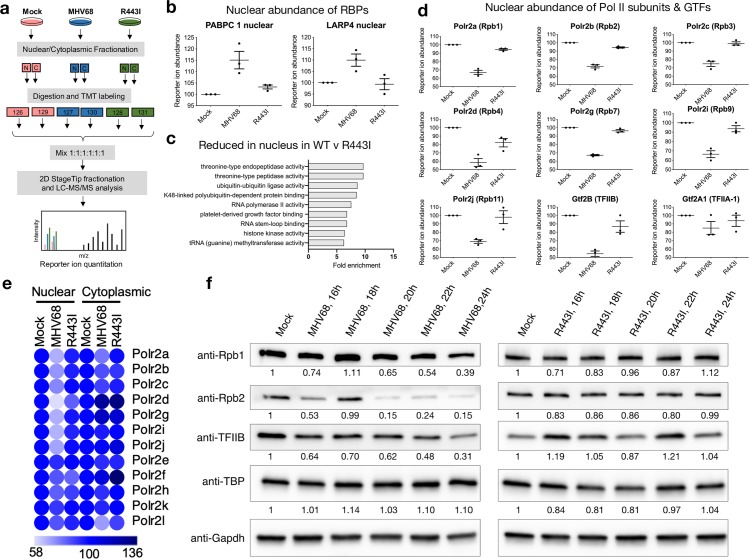
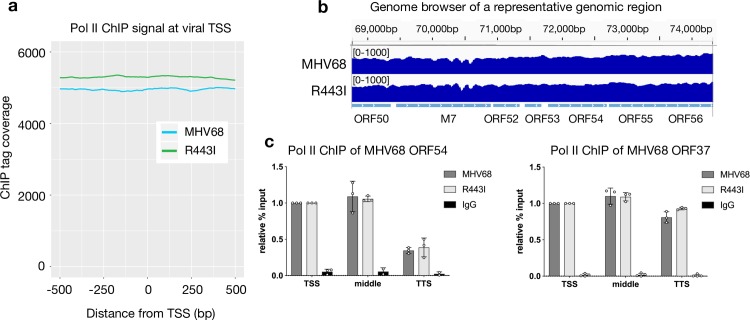
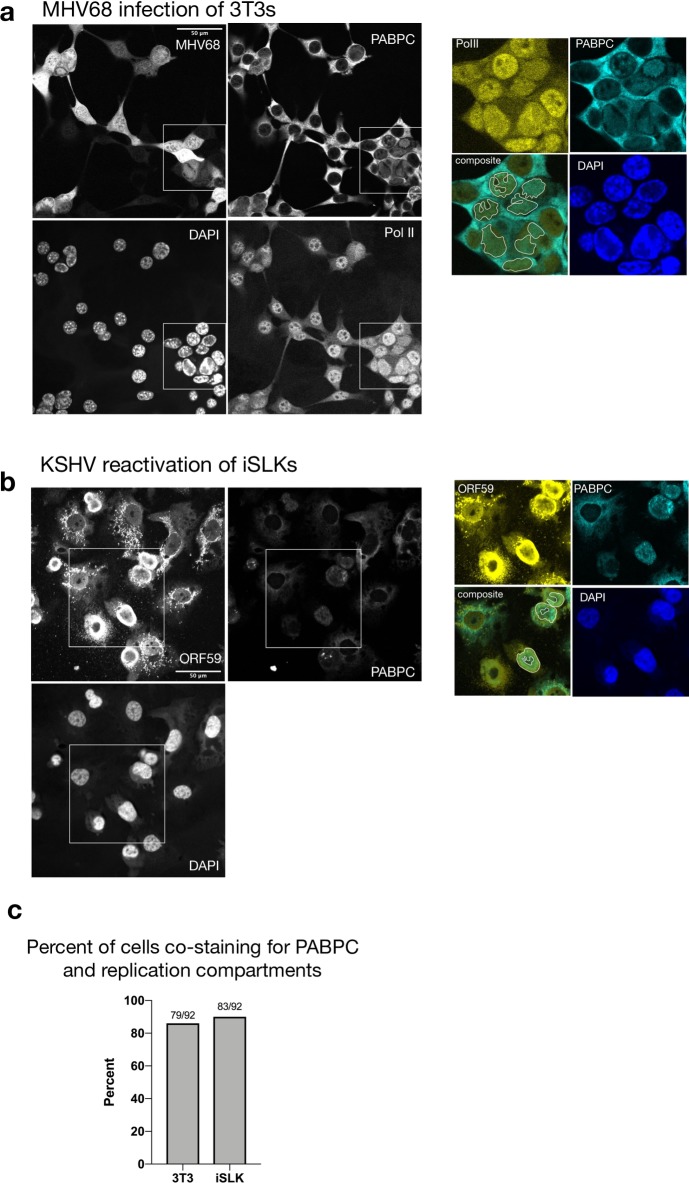
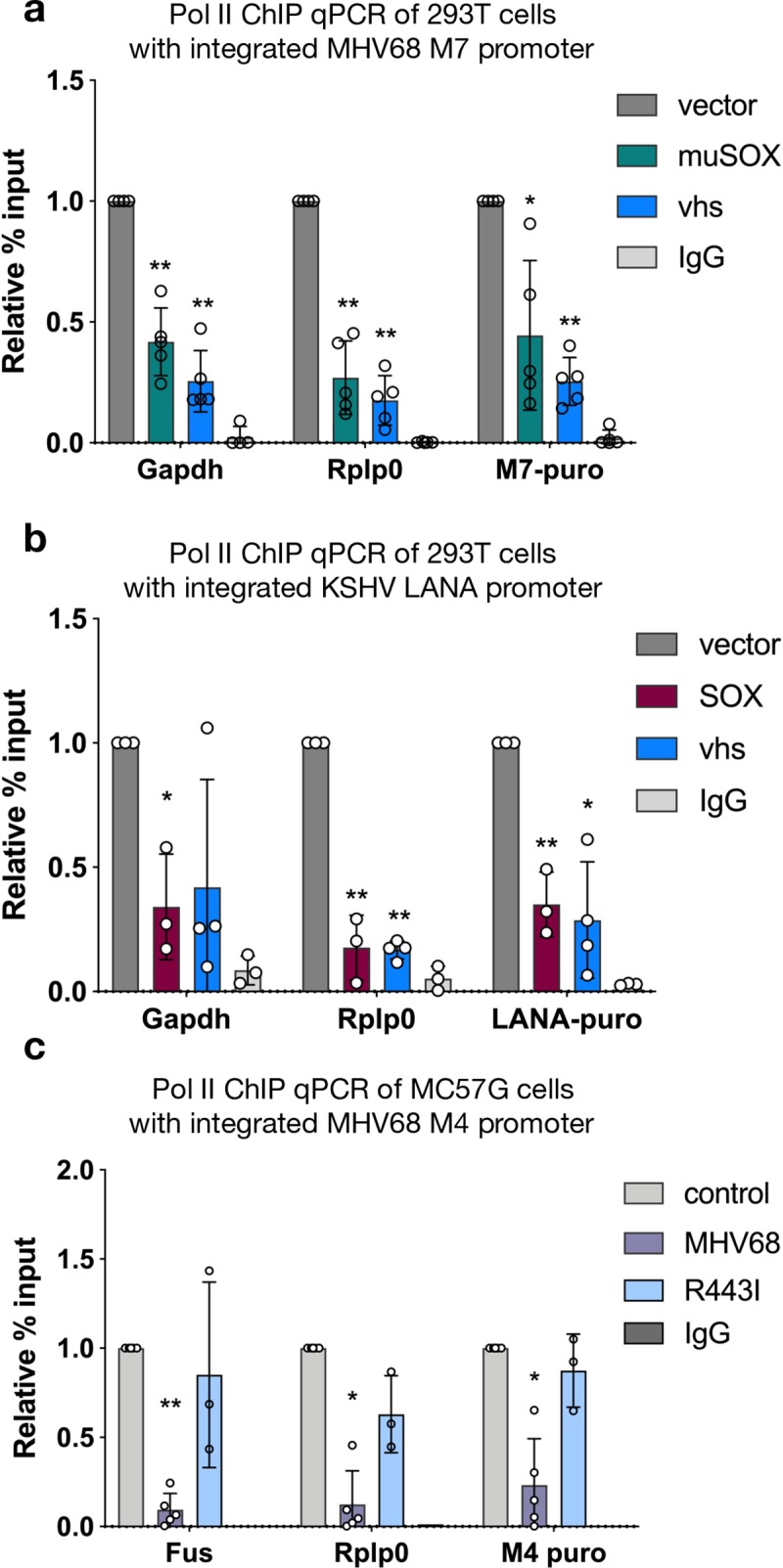
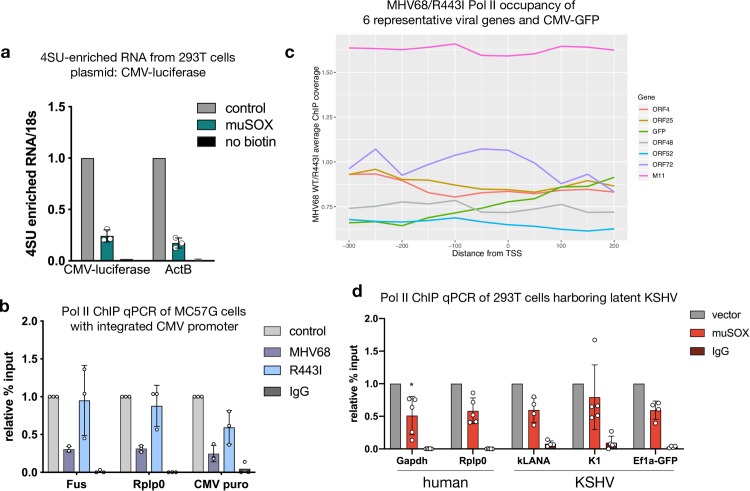
In mammalian cells, widespread acceleration of cytoplasmic mRNA degradation is linked to impaired RNA polymerase II (Pol II) transcription. This mRNA decay-induced transcriptional repression occurs during infection with gammaherpesviruses including Kaposi's sarcoma-associated herpesvirus (KSHV) and murine gammaherpesvirus 68 (MHV68), which encode an mRNA endonuclease that initiates widespread RNA decay. Here, we show that MHV68-induced mRNA decay leads to a genome-wide reduction of Pol II occupancy at mammalian promoters. This reduced Pol II occupancy is accompanied by down-regulation of multiple Pol II subunits and TFIIB in the nucleus of infected cells, as revealed by mass spectrometry-based global measurements of protein abundance. Viral genes, despite the fact that they require Pol II for transcription, escape transcriptional repression. Protection is not governed by viral promoter sequences; instead, location on the viral genome is both necessary and sufficient to escape the transcriptional repression effects of mRNA decay. We propose a model in which the ability to escape from transcriptional repression is linked to the localization of viral DNA within replication compartments, providing a means for these viruses to counteract decay-induced transcript loss.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures