

Posttranslational Modifications Mediate the Structural Diversity of Tauopathy Strains
- PMID: 32032505
- PMCID: PMC7491959
- DOI: 10.1016/j.cell.2020.01.027
Posttranslational Modifications Mediate the Structural Diversity of Tauopathy Strains
Erratum in
-
Posttranslational Modifications Mediate the Structural Diversity of Tauopathy Strains.Cell. 2021 Dec 9;184(25):6207-6210. doi: 10.1016/j.cell.2021.11.029. Cell. 2021. PMID: 34890553 Free PMC article. No abstract available.
Abstract
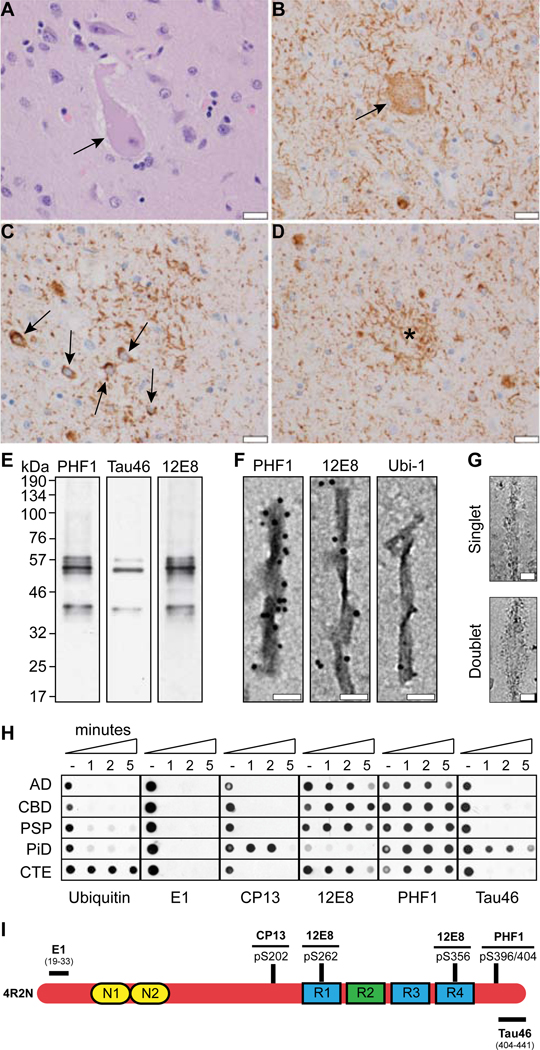
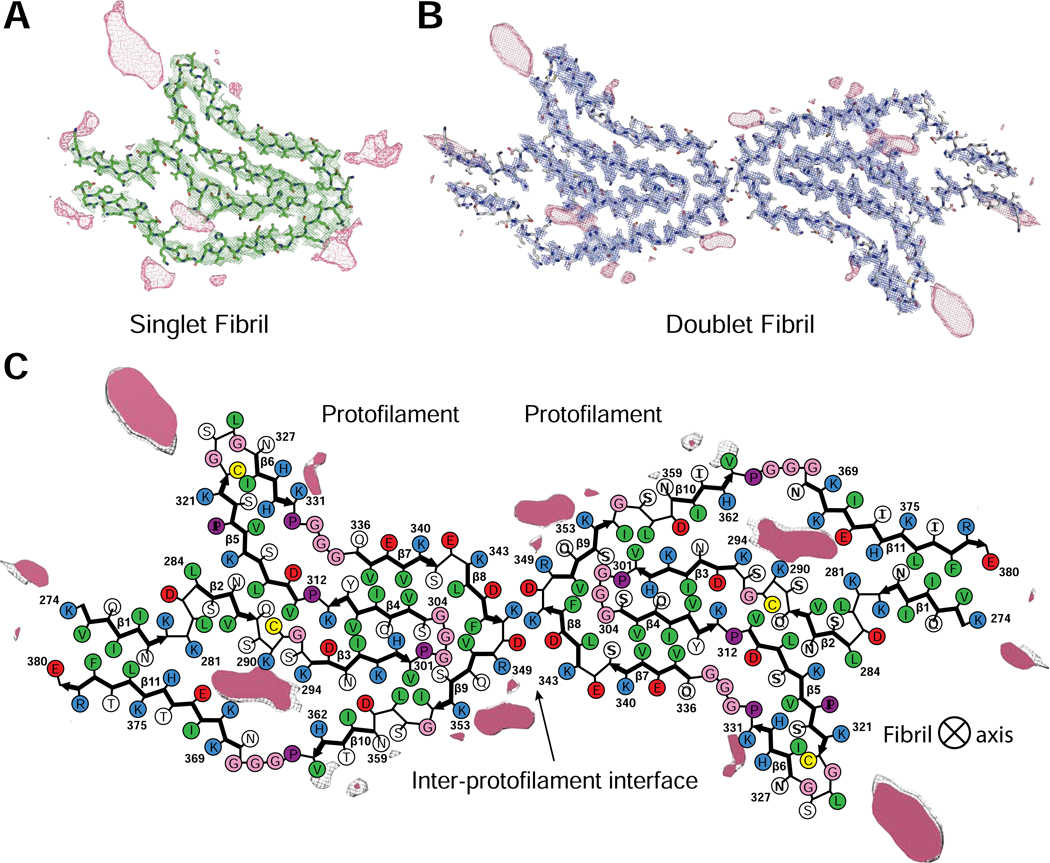
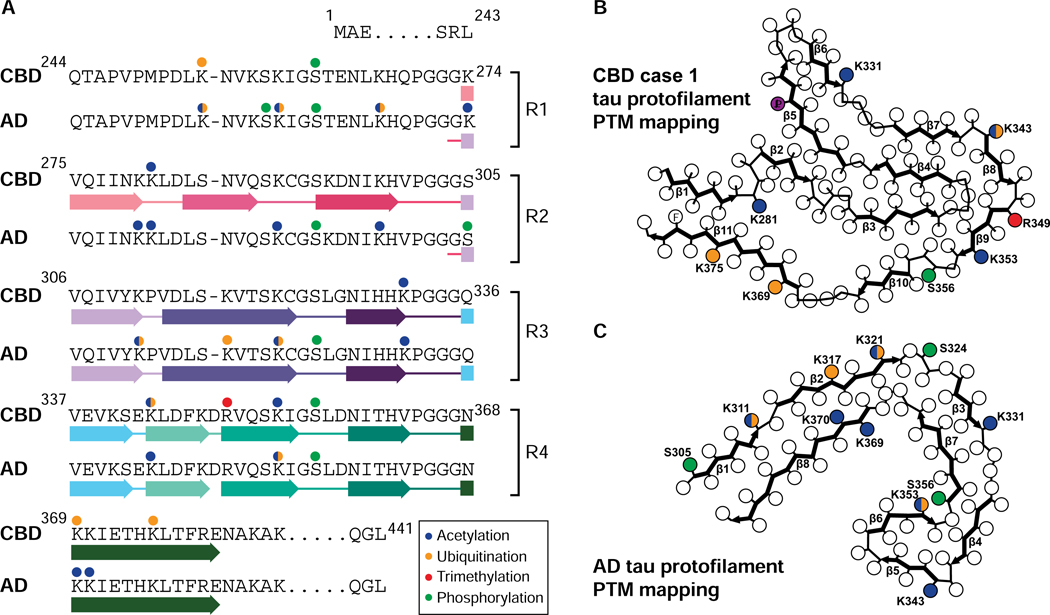
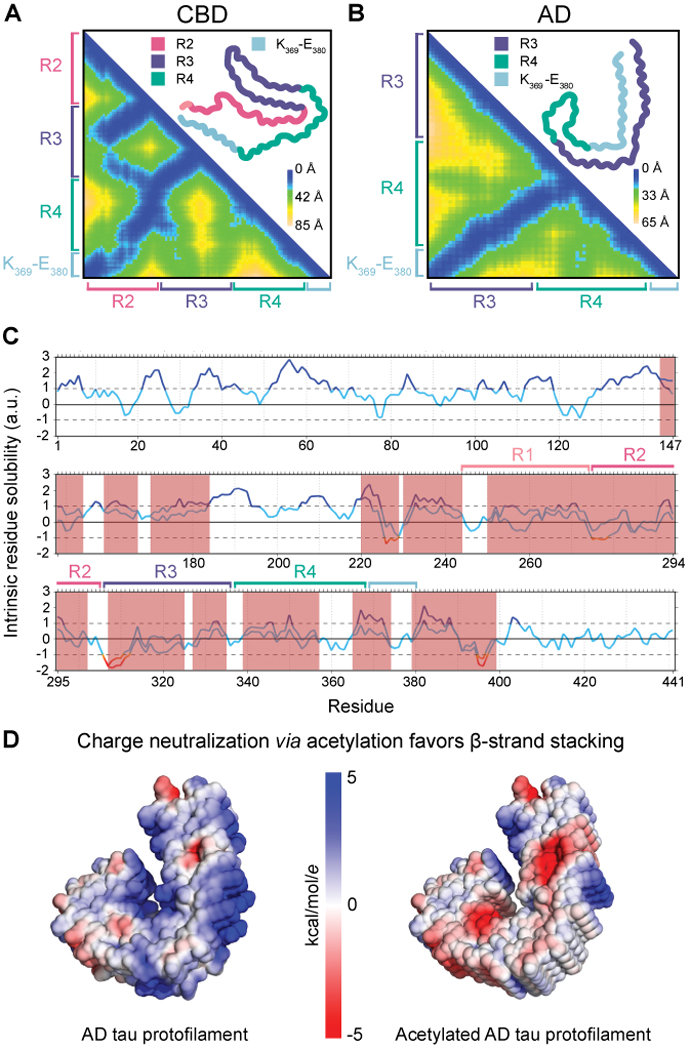
Tau aggregation into insoluble filaments is the defining pathological hallmark of tauopathies. However, it is not known what controls the formation and templated seeding of strain-specific structures associated with individual tauopathies. Here, we use cryo-electron microscopy (cryo-EM) to determine the structures of tau filaments from corticobasal degeneration (CBD) human brain tissue. Cryo-EM and mass spectrometry of tau filaments from CBD reveal that this conformer is heavily decorated with posttranslational modifications (PTMs), enabling us to map PTMs directly onto the structures. By comparing the structures and PTMs of tau filaments from CBD and Alzheimer's disease, it is found that ubiquitination of tau can mediate inter-protofilament interfaces. We propose a structure-based model in which cross-talk between PTMs influences tau filament structure, contributing to the structural diversity of tauopathy strains. Our approach establishes a framework for further elucidating the relationship between the structures of polymorphic fibrils, including their PTMs, and neurodegenerative disease.
Keywords: Alzheimer's disease; acetylation; corticobasal degeneration; cryo-EM; integrated structural biology; posttranslational modifications; tau strains; tauopathy; templated seeding; ubiquitination.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
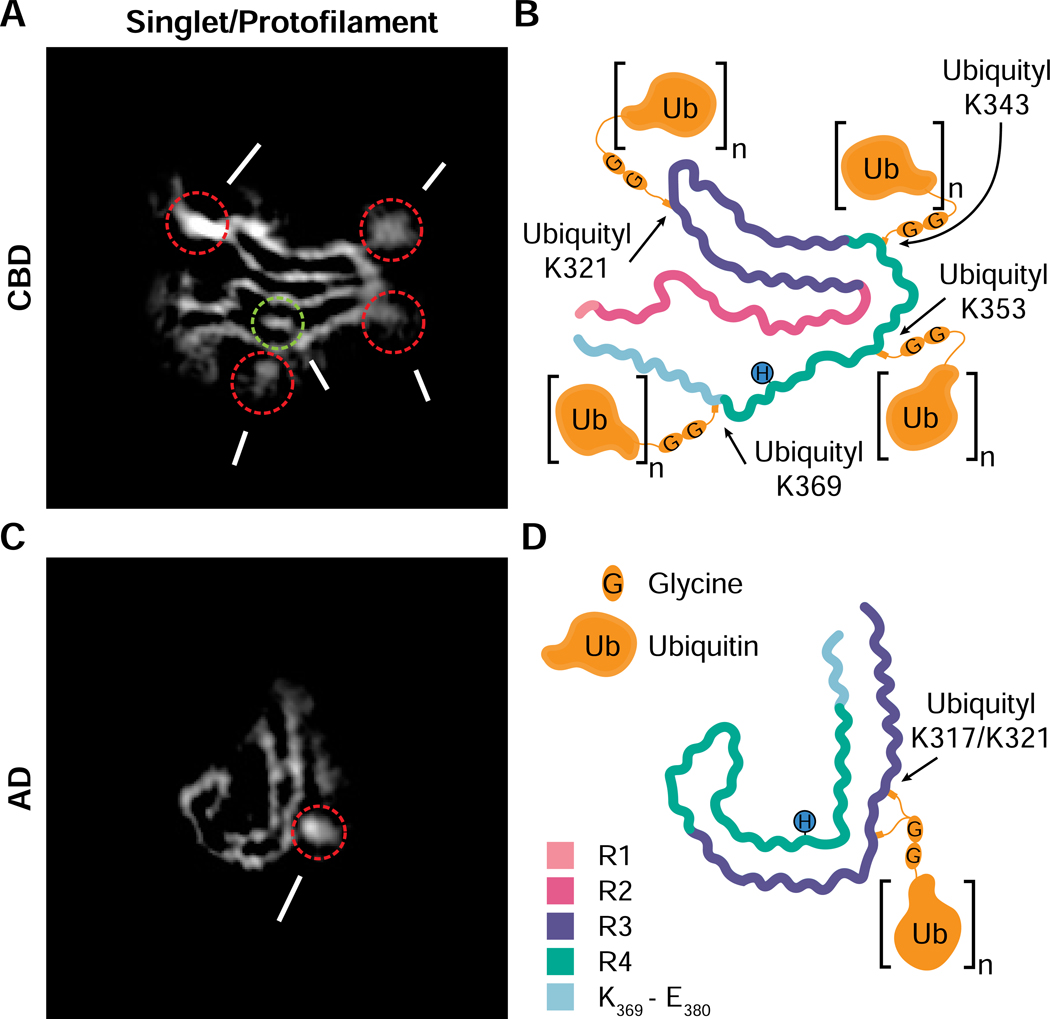
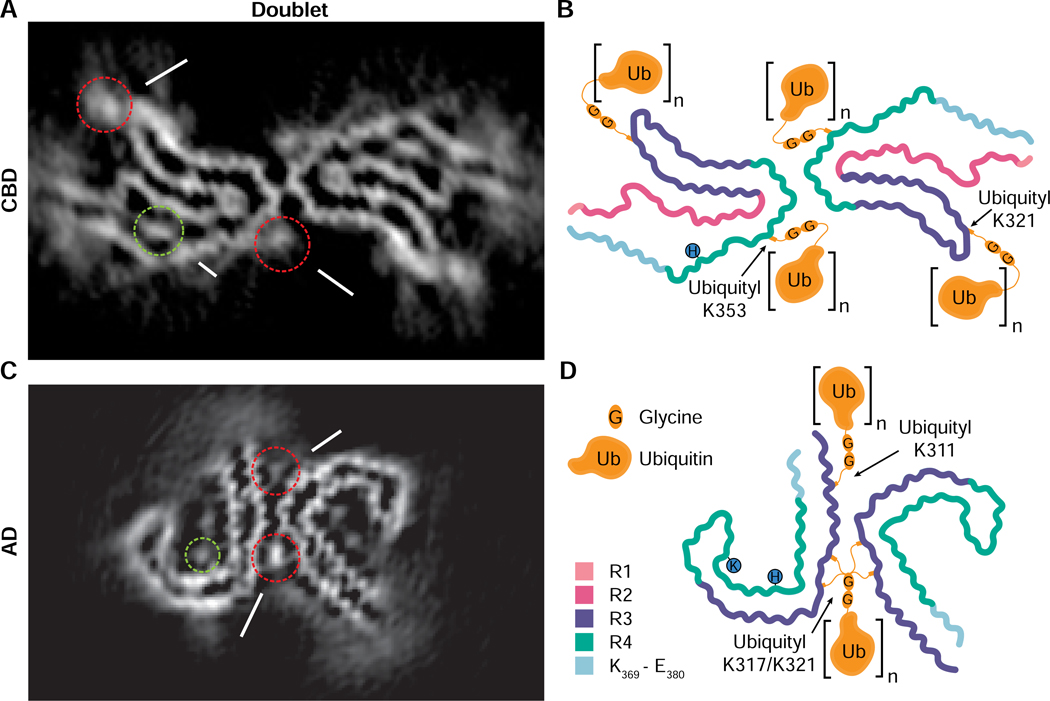
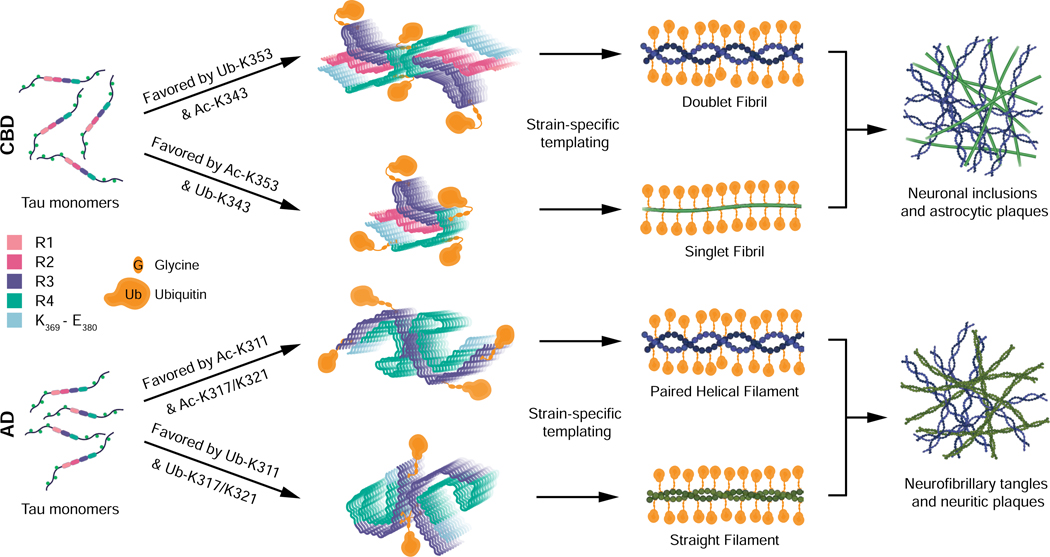
Comment in
-
Tau intricacies in disease.Nat Rev Mol Cell Biol. 2020 Apr;21(4):180. doi: 10.1038/s41580-020-0233-0. Nat Rev Mol Cell Biol. 2020. PMID: 32127686 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- R35 NS097273/NS/NINDS NIH HHS/United States
- RF1 AG062171/AG/NIA NIH HHS/United States
- R01 AG061800/AG/NIA NIH HHS/United States
- P41 GM103310/GM/NIGMS NIH HHS/United States
- P41 RR001209/RR/NCRR NIH HHS/United States
- U01 AG061357/AG/NIA NIH HHS/United States
- R01 NS088689/NS/NINDS NIH HHS/United States
- U24 GM116792/GM/NIGMS NIH HHS/United States
- RF1 AG056151/AG/NIA NIH HHS/United States
- P01 NS084974/NS/NINDS NIH HHS/United States
- S10 RR023057/RR/NCRR NIH HHS/United States
- U54 NS100693/NS/NINDS NIH HHS/United States
- U01 NS110438/NS/NINDS NIH HHS/United States
- R01 AG053960/AG/NIA NIH HHS/United States
- S10 OD018111/OD/NIH HHS/United States
- RF1 AG062077/AG/NIA NIH HHS/United States
- P01 NS099114/NS/NINDS NIH HHS/United States
- U01 AG046161/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
