

Impairment of Membrane Lipid Homeostasis by Bichalcone Analog TSWU-BR4 Attenuates Function of GRP78 in Regulation of the Oxidative Balance and Invasion of Cancer Cells
- PMID: 32033487
- PMCID: PMC7072528
- DOI: 10.3390/cells9020371
Impairment of Membrane Lipid Homeostasis by Bichalcone Analog TSWU-BR4 Attenuates Function of GRP78 in Regulation of the Oxidative Balance and Invasion of Cancer Cells
Abstract
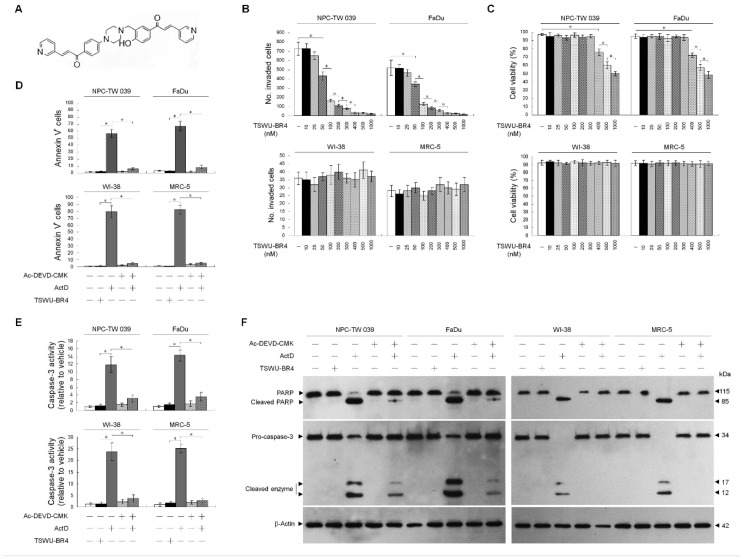
The specialized cholesterol/sphingolipid-rich membrane domains termed lipid rafts are highly dynamic in the cancer cells, which rapidly assemble effector molecules to form a sorting platform essential for oncogenic signaling transduction in response to extra- or intracellular stimuli. Density-based membrane flotation, subcellular fractionation, cell surface biotinylation, and co-immunoprecipitation analyses of bichalcone analog ((E)-1-(4-Hydroxy-3-((4-(4-((E)-3-(pyridin-3-yl)acryloyl)phenyl)piperazin-1-yl)methyl)phenyl)-3-(pyridin-3-yl)prop-2-en-1-one (TSWU-BR4)-treated cancer cells showed dissociation between GRP78 and p85α conferring the recruitment of PTEN to lipid raft membranes associated with p85α. Ectopic expression of GRP78 could overcome induction of lipid raft membrane-associated p85α-unphosphorylated PTEN complex formation and suppression of GRP78PI3KAktGTP-Rac1-mediated and GRP78-regulated PERKNrf2 antioxidant pathway and cancer cell invasion by TSWU-BR4. Using specific inducer, inhibitor, or short hairpin RNA for ASM demonstrated that induction of the lipid raft membrane localization and activation of ASM by TSWU-BR4 is responsible for perturbing homeostasis of cholesterol and ceramide levels in the lipid raft and ER membranes, leading to alteration of GRP78 membrane trafficking and subsequently inducing p85α-unphosphorylated PTEN complex formation, causing disruption of GRP78PI3KAktGTP-Rac1-mediated signal and ER membrane-associated GRP78-regulated oxidative stress balance, thus inhibiting cancer cell invasion. The involvement of the enrichment of ceramide to lipid raft membranes in inhibition of NF-κB-mediated MMP-2 expression was confirmed through attenuation of NF-κB activation using C2-ceramide, NF-κB specific inhibitors, ectopic expression of NF-κB p65, MMP-2 promoter-driven luciferase, and NF-κB-dependent reporter genes. In conclusion, localization of ASM in the lipid raft membranes by TSWU-BR4 is a key event for initiating formation of ceramide-enriched lipid raft membrane platforms, which causes delocalization of GRP78 from the lipid raft and ER membranes to the cytosol and formation of p85α-unphosphorylated PTEN complexes to attenuate the GRP78-regulated oxidative stress balance and GRP78p85αAktGTP-Rac1NF-κBMMP-2-mediated cancer cell invasion.
Keywords: ASM; GRP78; PTEN; ROS; bichalcone analog; ceramide; lipid raft; p85α.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
