

Increased circulating levels of Factor H-Related Protein 4 are strongly associated with age-related macular degeneration
- PMID: 32034129
- PMCID: PMC7005798
- DOI: 10.1038/s41467-020-14499-3
Increased circulating levels of Factor H-Related Protein 4 are strongly associated with age-related macular degeneration
Abstract
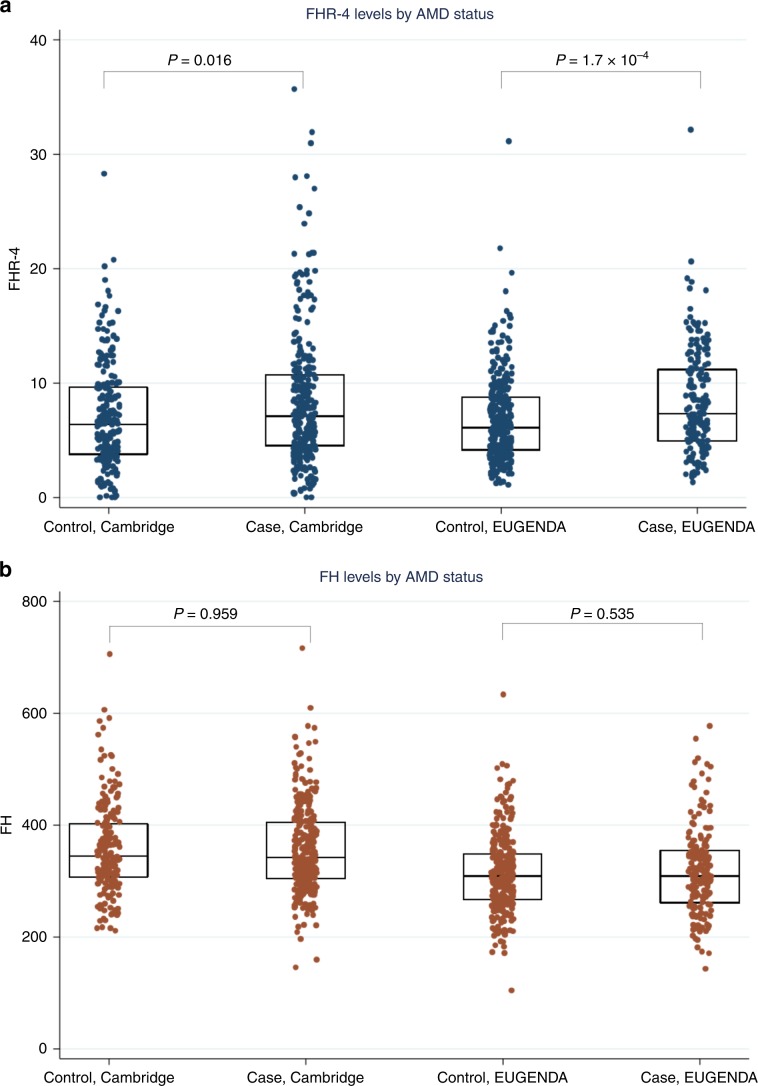
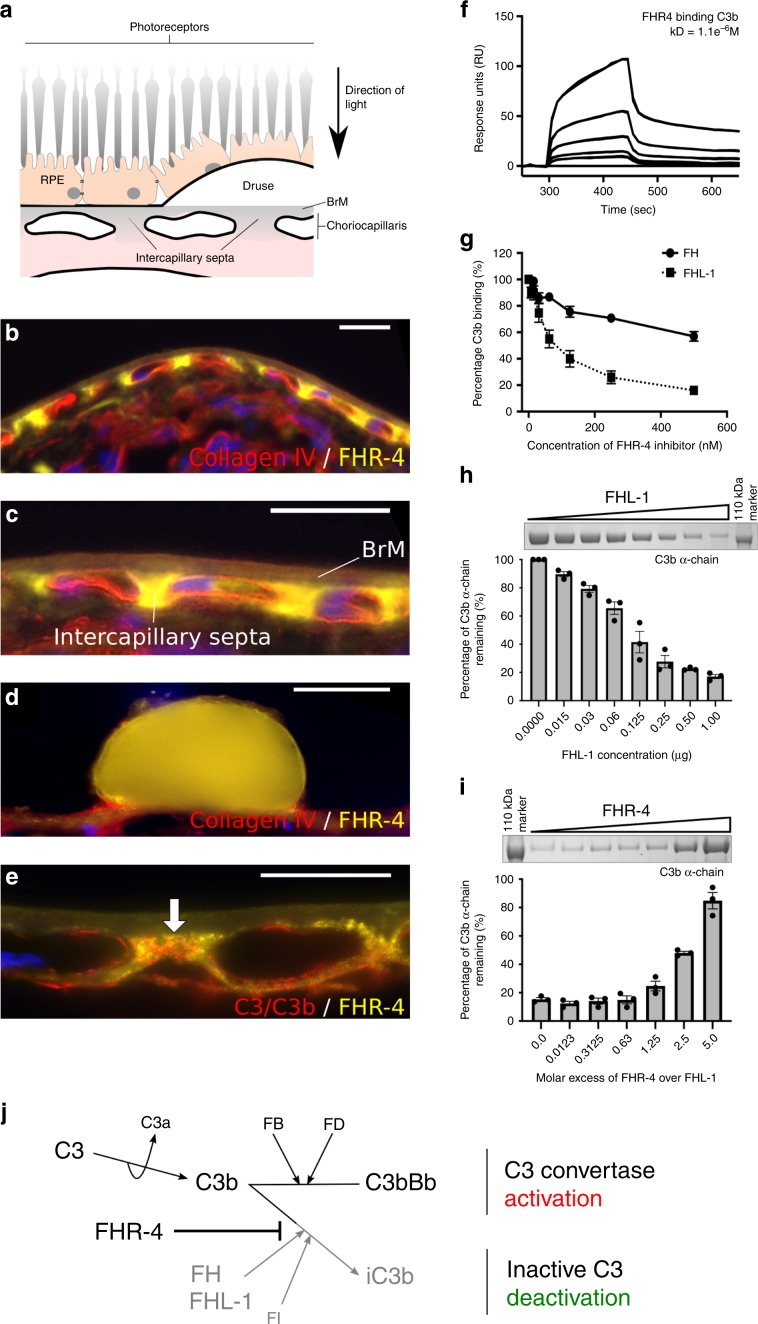
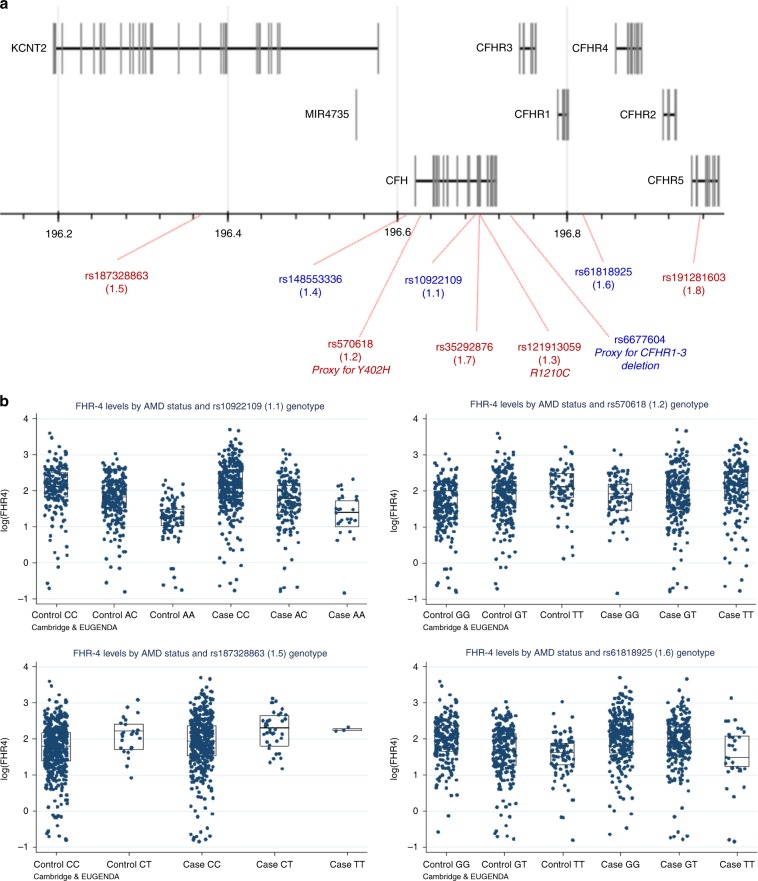
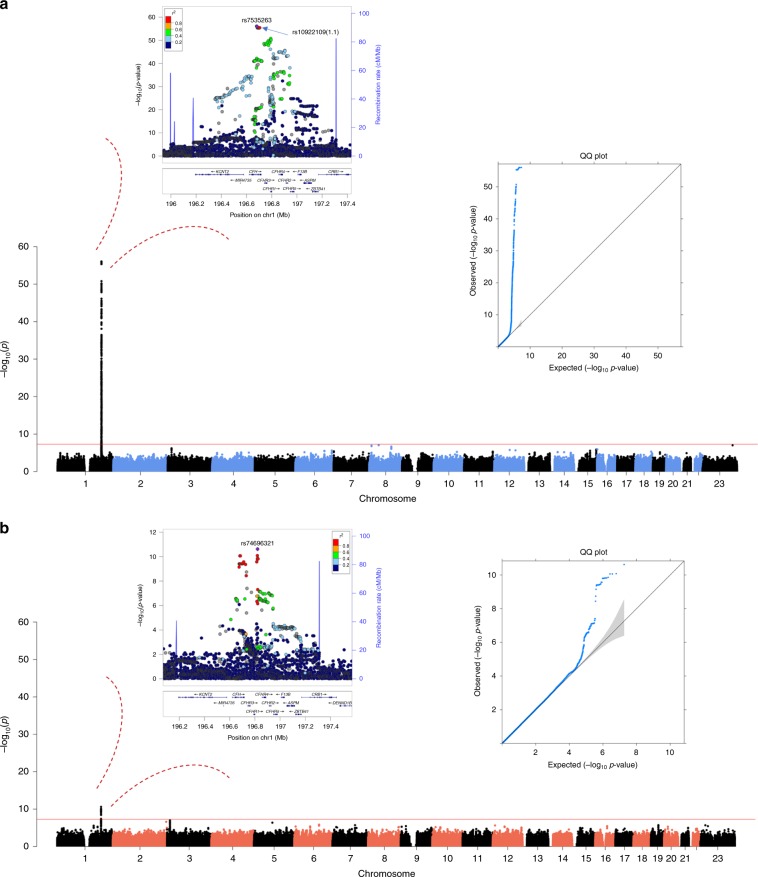
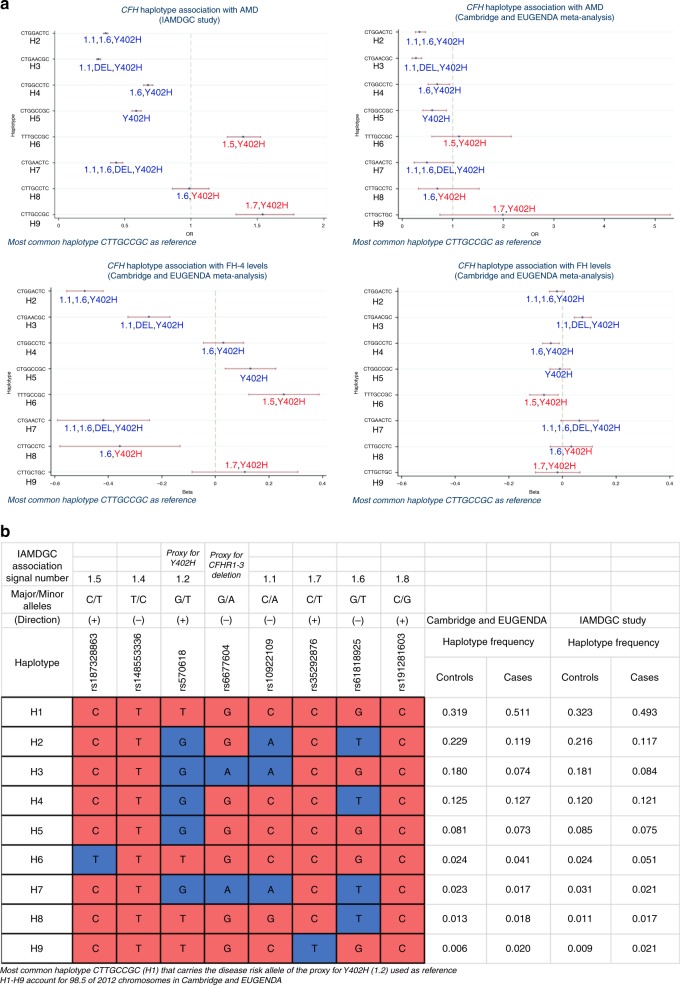
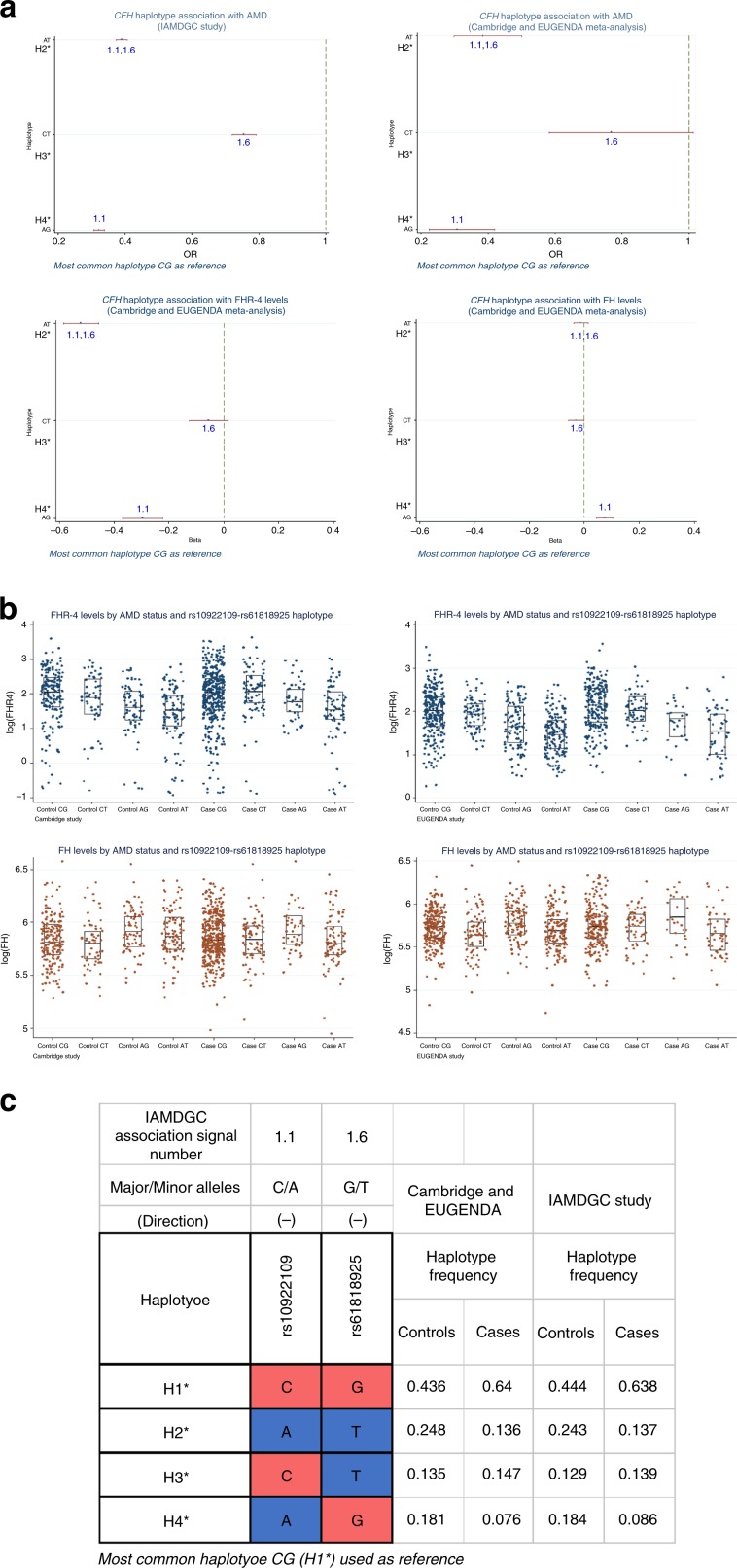
Age-related macular degeneration (AMD) is a leading cause of blindness. Genetic variants at the chromosome 1q31.3 encompassing the complement factor H (CFH, FH) and CFH related genes (CFHR1-5) are major determinants of AMD susceptibility, but their molecular consequences remain unclear. Here we demonstrate that FHR-4 plays a prominent role in AMD pathogenesis. We show that systemic FHR-4 levels are elevated in AMD (P-value = 7.1 × 10-6), whereas no difference is seen for FH. Furthermore, FHR-4 accumulates in the choriocapillaris, Bruch's membrane and drusen, and can compete with FH/FHL-1 for C3b binding, preventing FI-mediated C3b cleavage. Critically, the protective allele of the strongest AMD-associated CFH locus variant rs10922109 has the highest association with reduced FHR-4 levels (P-value = 2.2 × 10-56), independently of the AMD-protective CFHR1-3 deletion, and even in those individuals that carry the high-risk allele of rs1061170 (Y402H). Our findings identify FHR-4 as a key molecular player contributing to complement dysregulation in AMD.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
