

Cargo hold and delivery: Ankyrins, spectrins, and their functional patterning of neurons
- PMID: 32034889
- PMCID: PMC7705636
- DOI: 10.1002/cm.21602
Cargo hold and delivery: Ankyrins, spectrins, and their functional patterning of neurons
Abstract
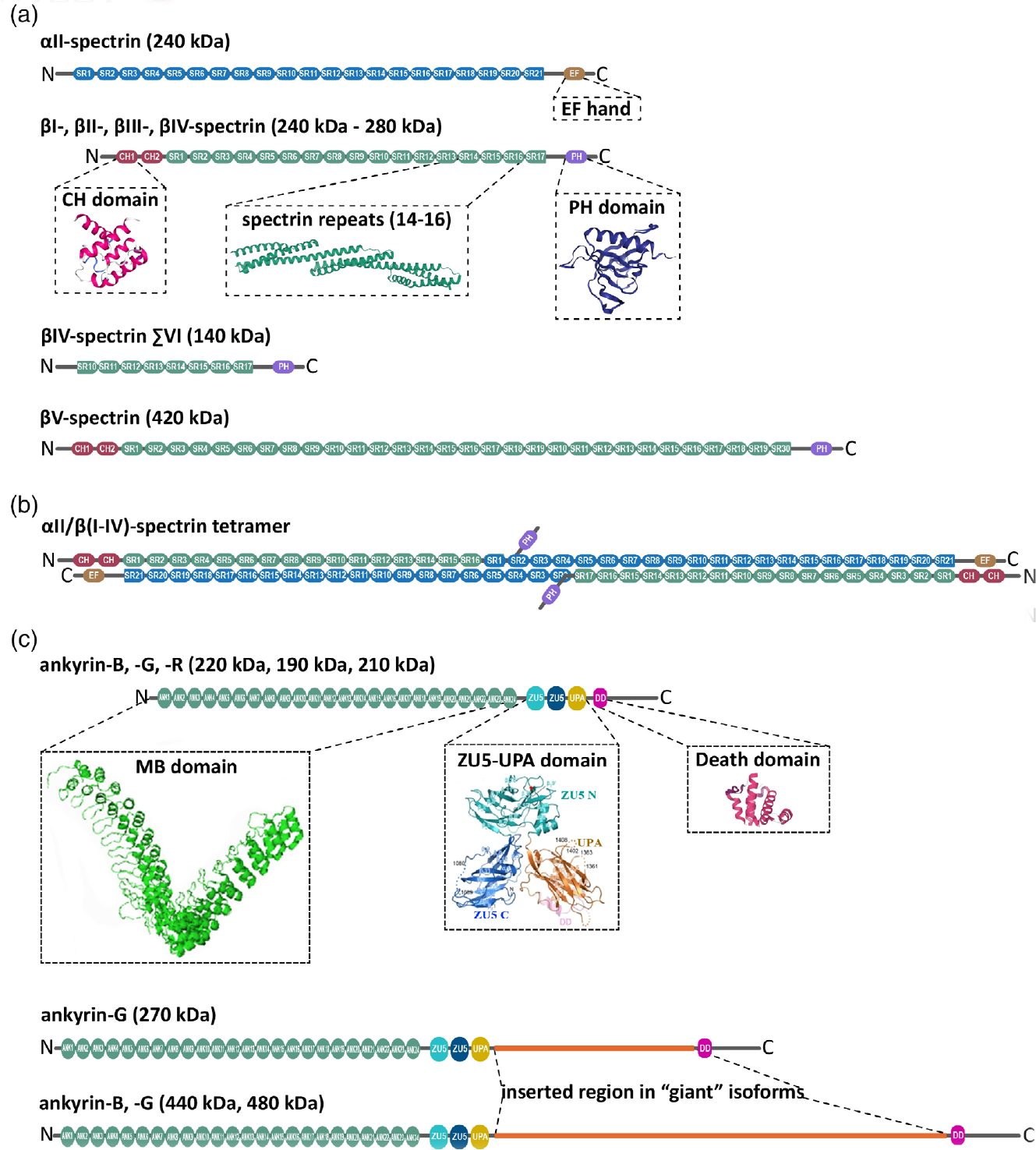
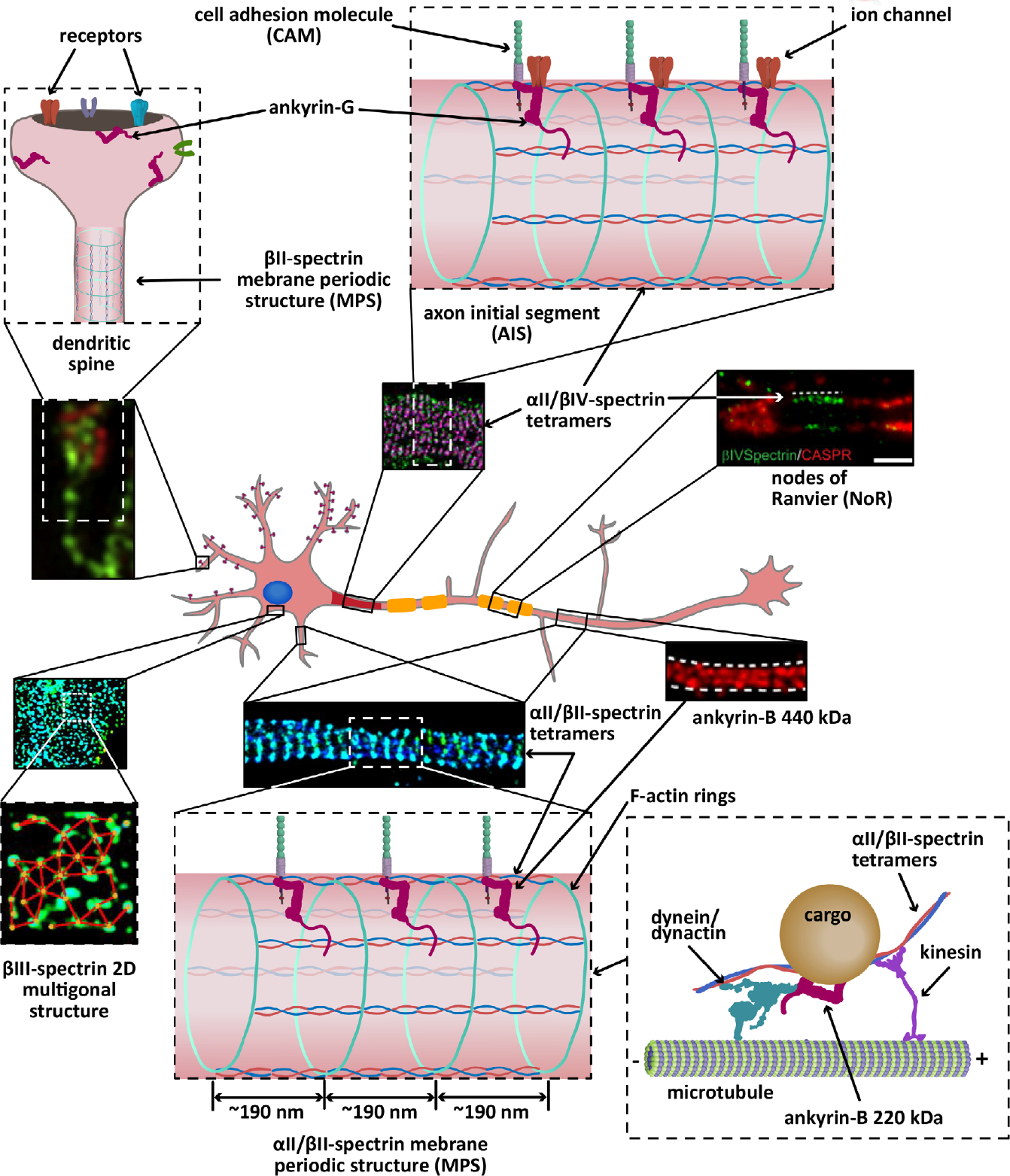
The highly polarized, typically very long, and nonmitotic nature of neurons present them with unique challenges in the maintenance of their homeostasis. This architectural complexity serves a rich and tightly controlled set of functions that enables their fast communication with neighboring cells and endows them with exquisite plasticity. The submembrane neuronal cytoskeleton occupies a pivotal position in orchestrating the structural patterning that determines local and long-range subcellular specialization, membrane dynamics, and a wide range of signaling events. At its center is the partnership between ankyrins and spectrins, which self-assemble with both remarkable long-range regularity and micro- and nanoscale specificity to precisely position and stabilize cell adhesion molecules, membrane transporters, ion channels, and other cytoskeletal proteins. To accomplish these generally conserved, but often functionally divergent and spatially diverse, roles these partners use a combinatorial program of a couple of dozens interacting family members, whose code is not fully unraveled. In a departure from their scaffolding roles, ankyrins and spectrins also enable the delivery of material to the plasma membrane by facilitating intracellular transport. Thus, it is unsurprising that deficits in ankyrins and spectrins underlie several neurodevelopmental, neurodegenerative, and psychiatric disorders. Here, I summarize key aspects of the biology of spectrins and ankyrins in the mammalian neuron and provide a snapshot of the latest advances in decoding their roles in the nervous system.
Keywords: ankyrin; axonal transport; cytoskeleton; plasma membrane; spectrin.
© 2020 Wiley Periodicals, Inc.
Figures
References
-
- Alemu EA, Lamark T, Torgersen KM, Birgisdottir AB, Larsen KB, Jain A, et al. (2012). ATG8 family proteins act as scaffolds for assembly of the ULK complex: Sequence requirements for LC3-interacting region (LIR) motifs. Journal of Biological Chemistry, 287, 39275–39290. 10.1074/jbc.M112.378109 - DOI - PMC - PubMed
-
- An X, Guo X, Sum H, Morrow J, Gratzer W, & Mohandas N (2004). Phosphatidylserine binding sites in erythroid spectrin: Location and implications for membrane stability. Biochemistry, 43, 310–315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
