

Modality-specific attention attenuates visual-tactile integration and recalibration effects by reducing prior expectations of a common source for vision and touch
- PMID: 32036027
- PMCID: PMC7182122
- DOI: 10.1016/j.cognition.2019.104170
Modality-specific attention attenuates visual-tactile integration and recalibration effects by reducing prior expectations of a common source for vision and touch
Abstract
At any moment in time, streams of information reach the brain through the different senses. Given this wealth of noisy information, it is essential that we select information of relevance - a function fulfilled by attention - and infer its causal structure to eventually take advantage of redundancies across the senses. Yet, the role of selective attention during causal inference in cross-modal perception is unknown. We tested experimentally whether the distribution of attention across vision and touch enhances cross-modal spatial integration (visual-tactile ventriloquism effect, Expt. 1) and recalibration (visual-tactile ventriloquism aftereffect, Expt. 2) compared to modality-specific attention, and then used causal-inference modeling to isolate the mechanisms behind the attentional modulation. In both experiments, we found stronger effects of vision on touch under distributed than under modality-specific attention. Model comparison confirmed that participants used Bayes-optimal causal inference to localize visual and tactile stimuli presented as part of a visual-tactile stimulus pair, whereas simultaneously collected unity judgments - indicating whether the visual-tactile pair was perceived as spatially-aligned - relied on a sub-optimal heuristic. The best-fitting model revealed that attention modulated sensory and cognitive components of causal inference. First, distributed attention led to an increase of sensory noise compared to selective attention toward one modality. Second, attending to both modalities strengthened the stimulus-independent expectation that the two signals belong together, the prior probability of a common source for vision and touch. Yet, only the increase in the expectation of vision and touch sharing a common source was able to explain the observed enhancement of visual-tactile integration and recalibration effects with distributed attention. In contrast, the change in sensory noise explained only a fraction of the observed enhancements, as its consequences vary with the overall level of noise and stimulus congruency. Increased sensory noise leads to enhanced integration effects for visual-tactile pairs with a large spatial discrepancy, but reduced integration effects for stimuli with a small or no cross-modal discrepancy. In sum, our study indicates a weak a priori association between visual and tactile spatial signals that can be strengthened by distributing attention across both modalities.
Keywords: Attention; Causal inference; Multisensory integration; Recalibration; Ventriloquism; Visual-tactile.
Copyright © 2019 Elsevier B.V. All rights reserved.
Figures



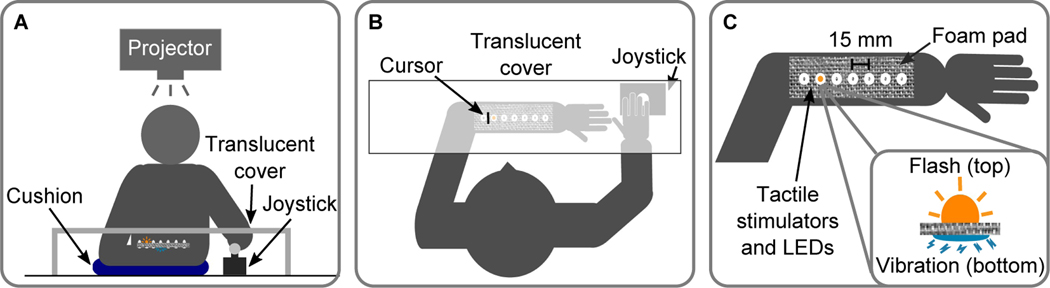
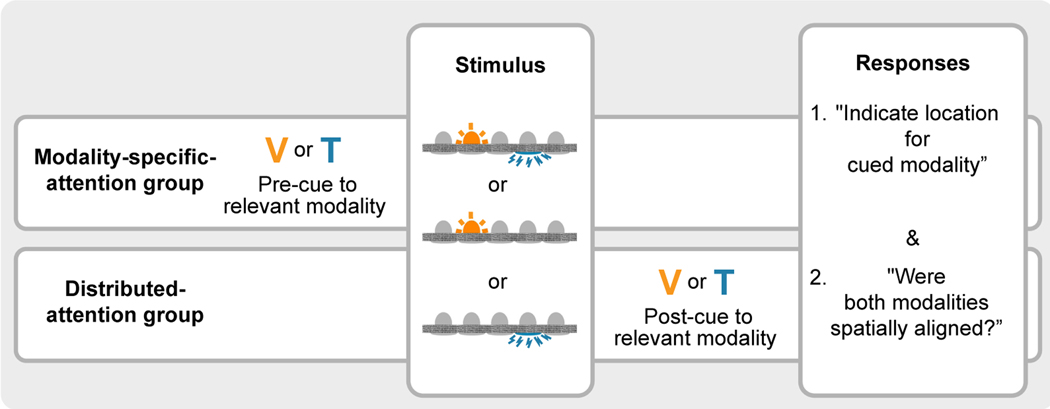
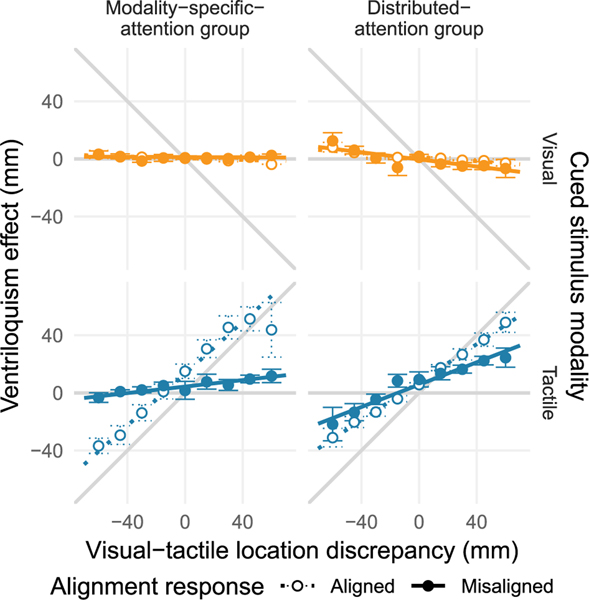


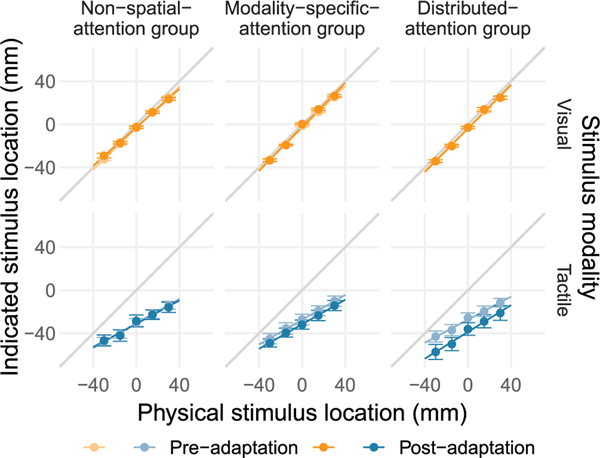





References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
