

Dihydrotestosterone Regulates Hair Growth Through the Wnt/β-Catenin Pathway in C57BL/6 Mice and In Vitro Organ Culture
- PMID: 32038233
- PMCID: PMC6989660
- DOI: 10.3389/fphar.2019.01528
Dihydrotestosterone Regulates Hair Growth Through the Wnt/β-Catenin Pathway in C57BL/6 Mice and In Vitro Organ Culture
Abstract
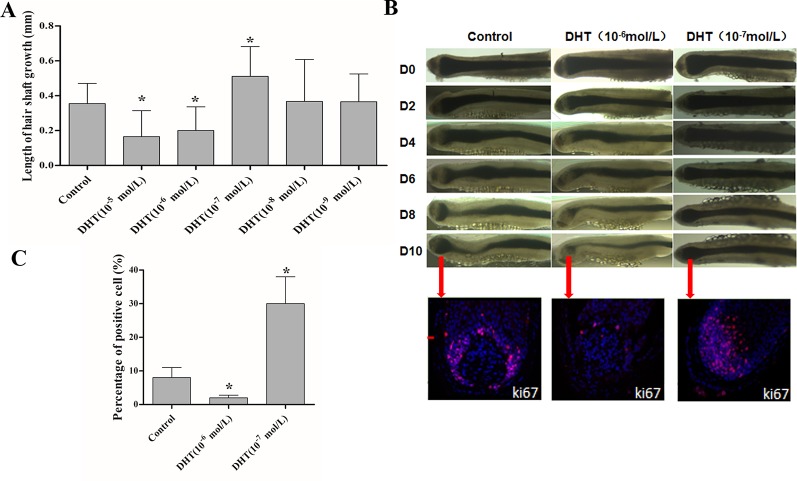
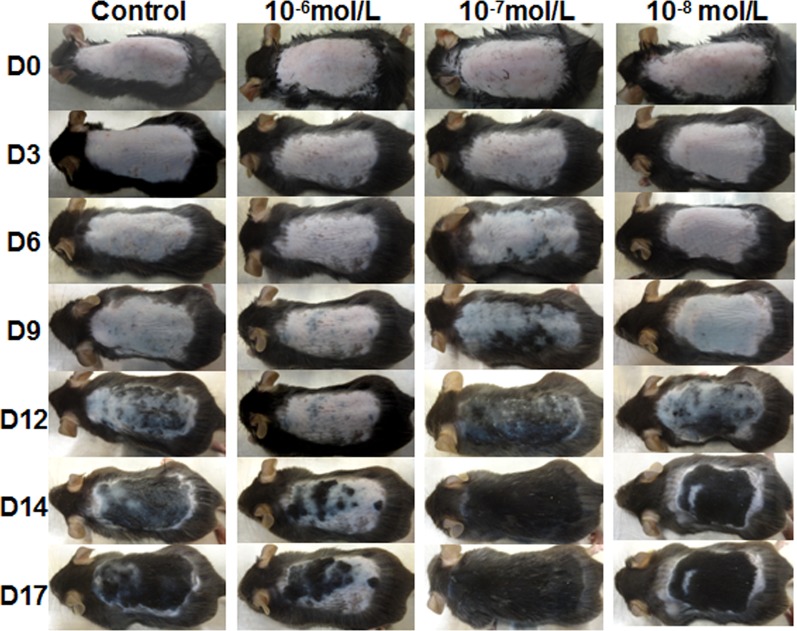
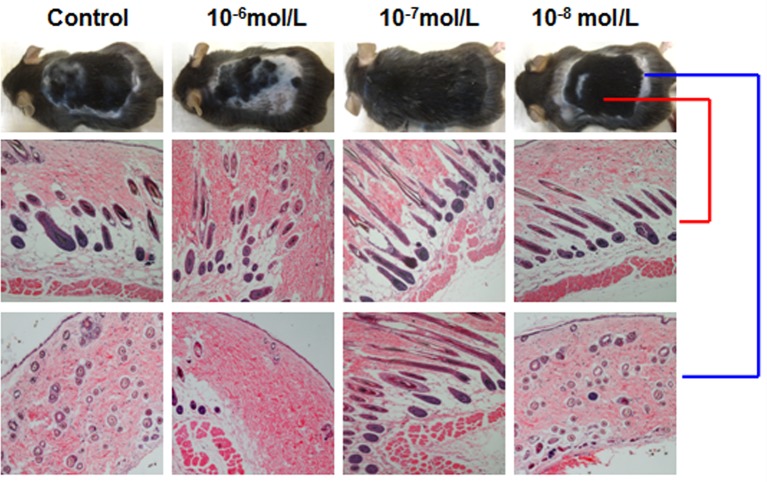
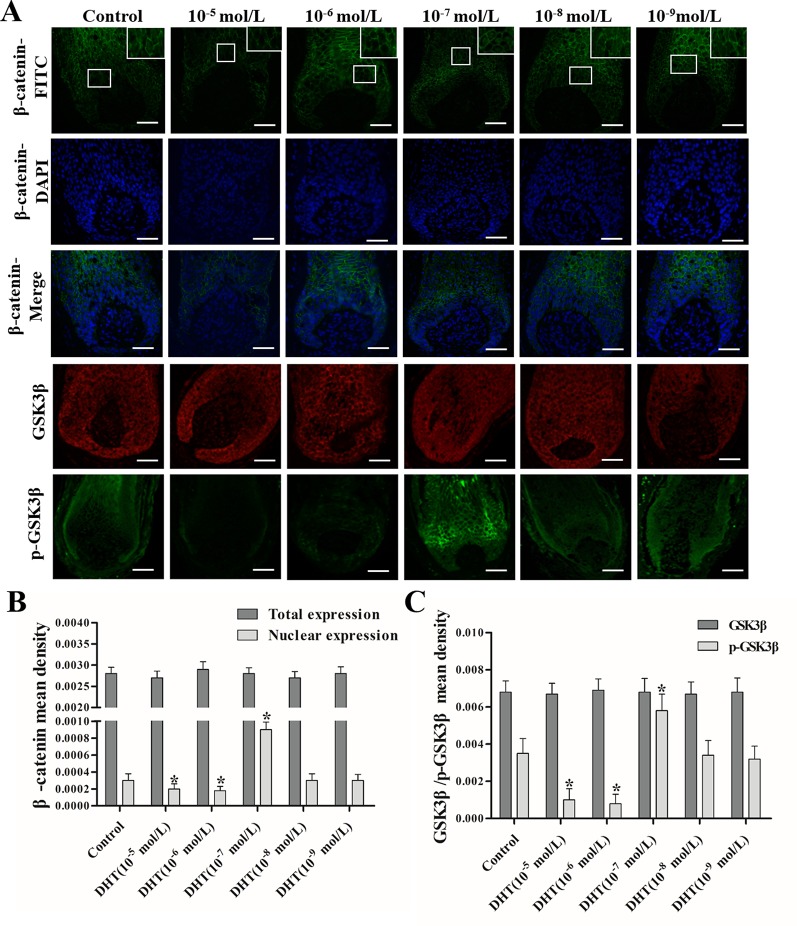
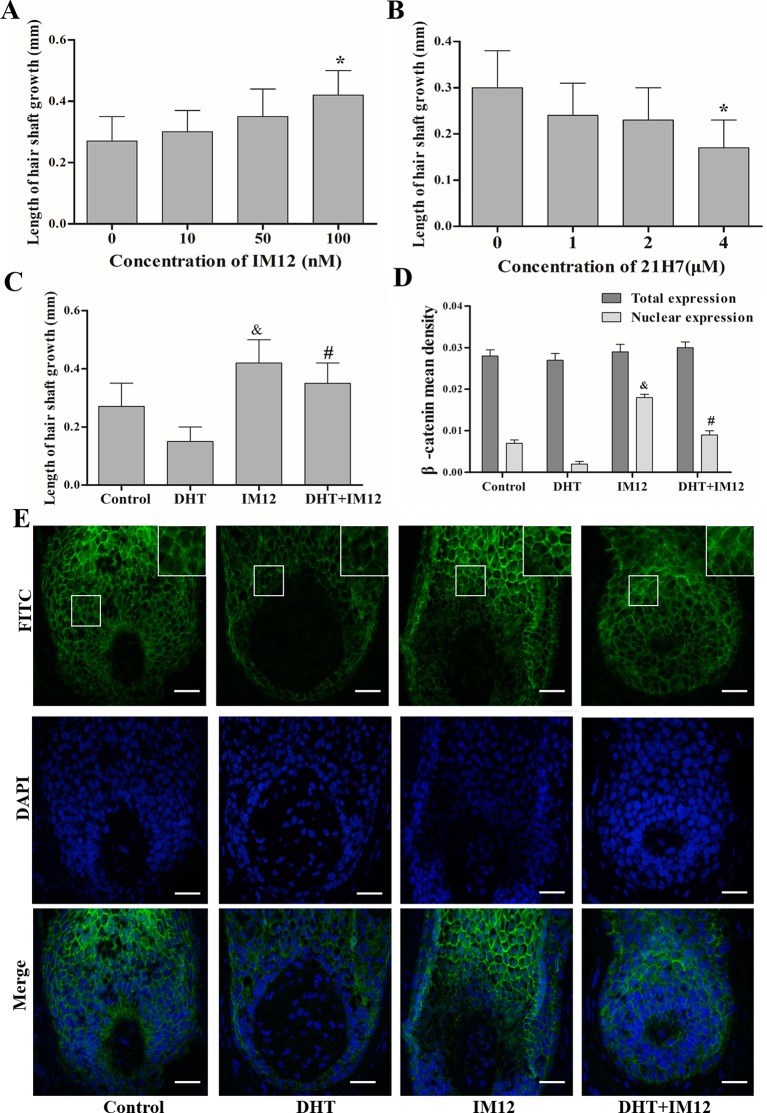
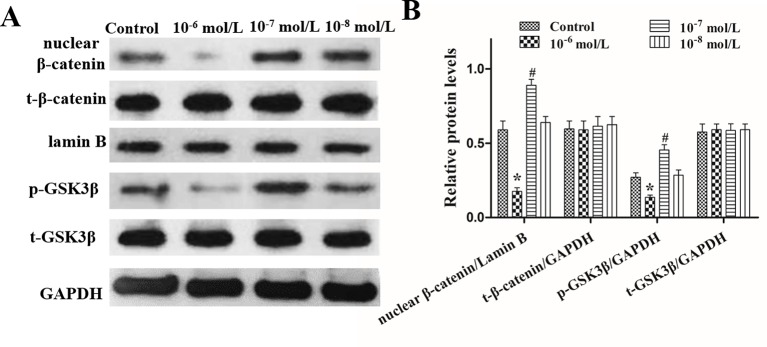
Dihydrotestosterone (DHT) is the most potent androgen that regulates hair cycling. Hair cycling involves cross-talk between the androgen and Wnt/β-catenin pathways. However, how DHT regulates hair follicle (HF) growth through the Wnt/β-catenin pathway has not been well investigated. This study aimed to investigate the roles of DHT in hair growth in vivo and in vitro. Human scalp HFs were treated with different concentrations of DHT (10-5, 10-6, 10-7, 10-8, and 10-9 mol/L) for 10 days. The effects of DHT on hair shaft elongation, the proliferation of hair matrix cells, and the levels of β-catenin, GSK-3β, and phosphorylated GSK-3β (ser9) were evaluated in the cultured HFs. The effects of DHT were further investigated in C57BL/6 mice. Moreover, the growth of cultured human HFs was observed after interfering with the β-catenin pathway through inhibitors or activators in the presence or absence of DHT. We found that different concentrations of DHT had different effects on human HFs in vitro and C57BL/6 mice. At 10-6 mol/L, DHT inhibited HF growth and β-catenin/p-GSK-3β expression, whereas 10-7 mol/L DHT induced HF growth and β-catenin/p-GSK-3β expression. In addition, a β-catenin inhibitor (21H7) inhibited HF growth in vitro, while a β-catenin activator (IM12) promoted HF growth in vitro and antagonized the inhibition of HFs by high levels of DHT. These results suggest that DHT plays a pivotal role in region-specific hair growth, which may be related to the Wnt/β-catenin pathway.
Keywords: Wnt/β-catenin pathway; androgenetic alopecia; dihydrotestosterone; hair follicle; hair growth; β-catenin activator.
Copyright © 2020 Chen, Liu, Li, Han, Tang, Deng, Lai and Wan.
Figures
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous