

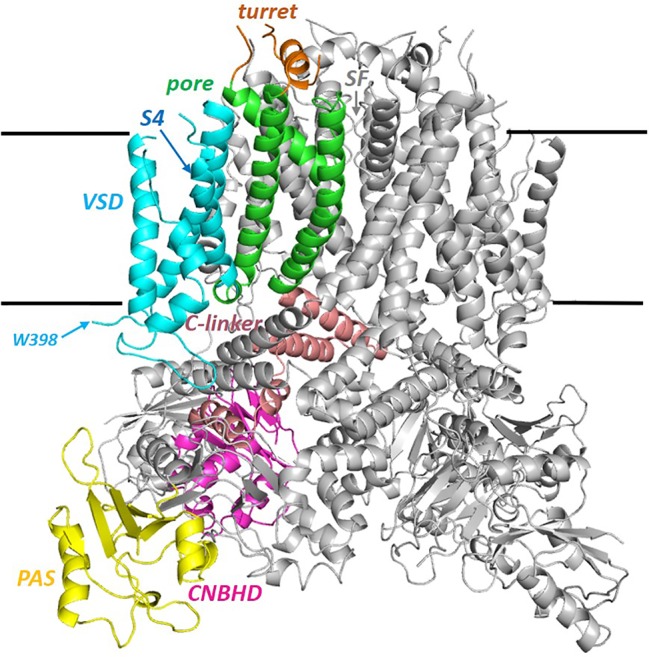
An Update on the Structure of hERG
- PMID: 32038248
- PMCID: PMC6992539
- DOI: 10.3389/fphar.2019.01572
An Update on the Structure of hERG
Abstract
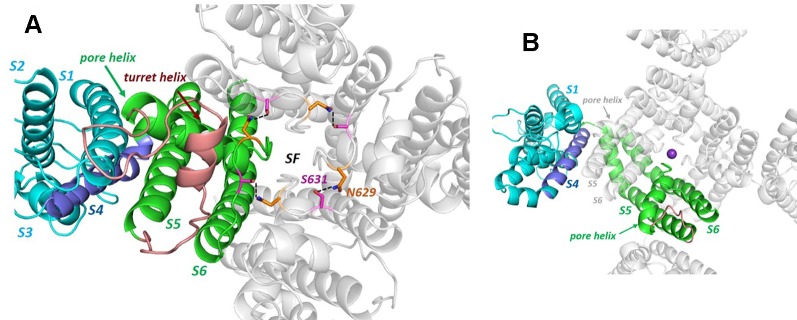
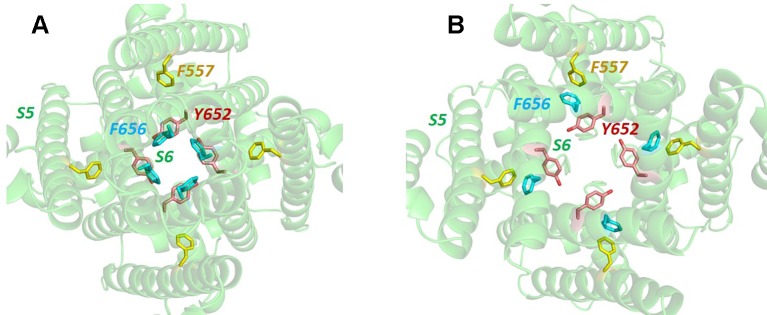
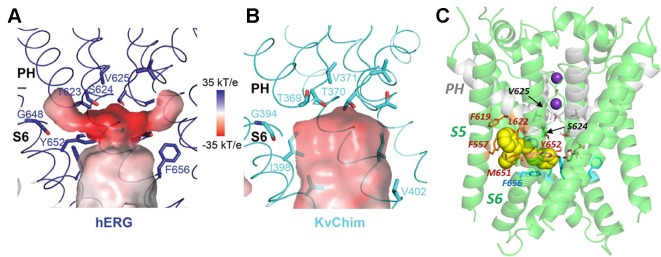
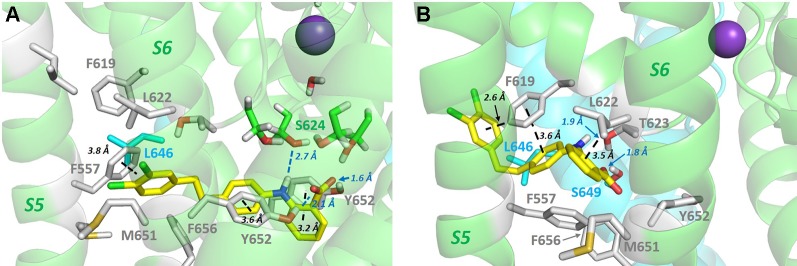
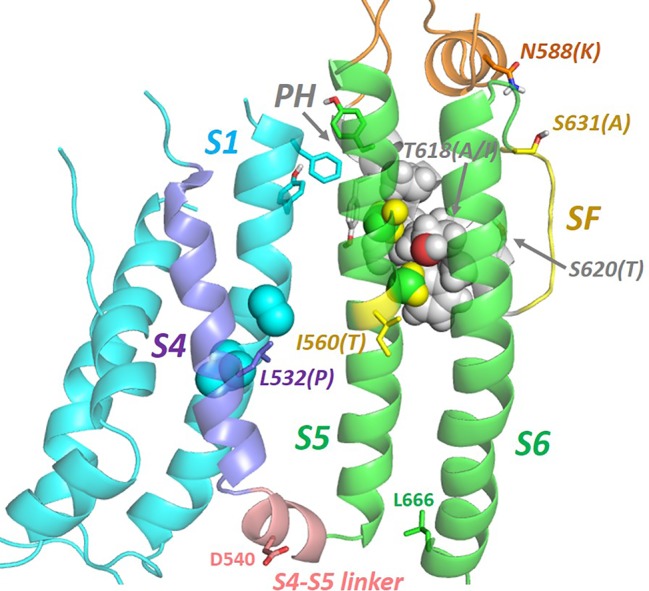
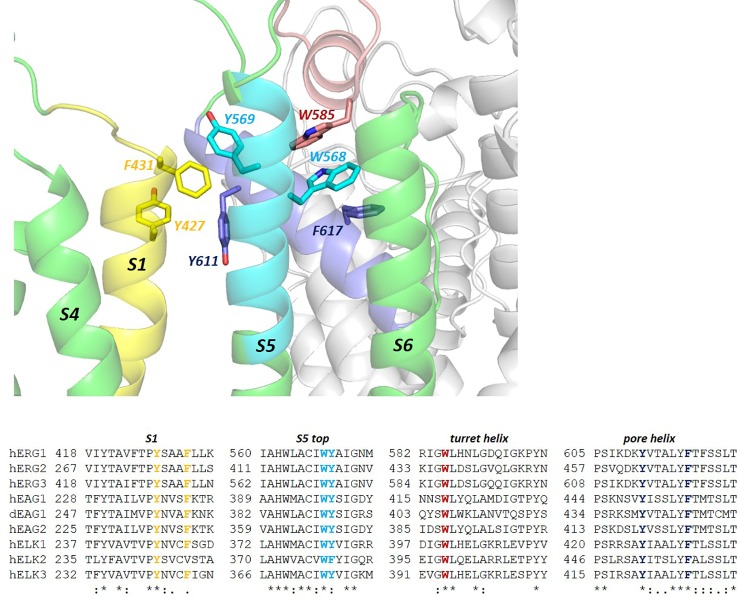
The human voltage-sensitive K+ channel hERG plays a fundamental role in cardiac action potential repolarization, effectively controlling the QT interval of the electrocardiogram. Inherited loss- or gain-of-function mutations in hERG can result in dangerous "long" (LQTS) or "short" QT syndromes (SQTS), respectively, and the anomalous susceptibility of hERG to block by a diverse range of drugs underlies an acquired LQTS. A recent open channel cryo-EM structure of hERG should greatly advance understanding of the molecular basis of hERG channelopathies and drug-induced LQTS. Here we describe an update of recent research that addresses the nature of the particular gated state of hERG captured in the new structure, and the insight afforded by the structure into the molecular basis for high affinity drug block of hERG, the binding of hERG activators and the molecular basis of hERG's peculiar gating properties. Interpretation of the pharmacology of natural SQTS mutants in the context of the structure is a promising approach to understanding the molecular basis of hERG inactivation, and the structure suggests how voltage-dependent changes in the membrane domain may be transmitted to an extracellular "turret" to effect inactivation through aromatic side chain motifs that are conserved throughout the KCNH family of channels.
Keywords: C-type inactivation; KCNH; channelopathy; cryo-EM structure; drug block; hERG; long QT syndrome; short QT syndrome.
Copyright © 2020 Butler, Helliwell, Zhang, Hancox and Dempsey.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources