

Whey Peptides Stimulate Differentiation and Lipid Metabolism in Adipocytes and Ameliorate Lipotoxicity-Induced Insulin Resistance in Muscle Cells
- PMID: 32041341
- PMCID: PMC7071342
- DOI: 10.3390/nu12020425
Whey Peptides Stimulate Differentiation and Lipid Metabolism in Adipocytes and Ameliorate Lipotoxicity-Induced Insulin Resistance in Muscle Cells
Abstract
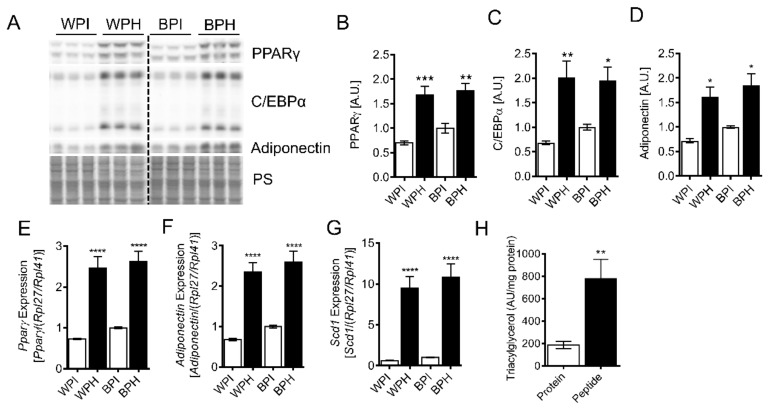
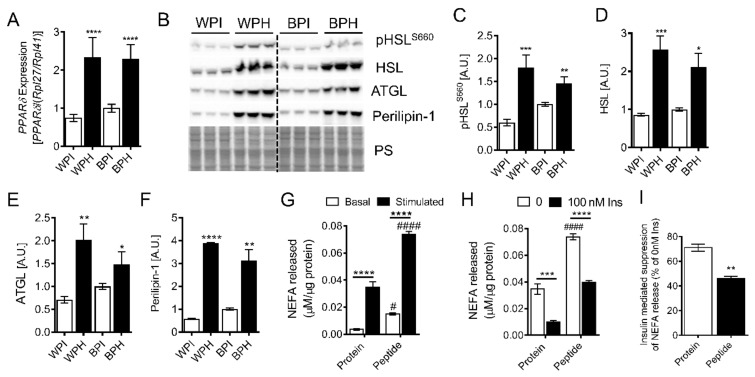
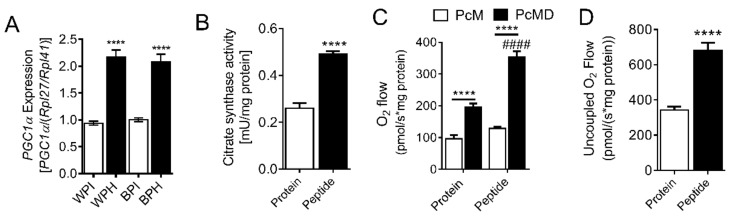
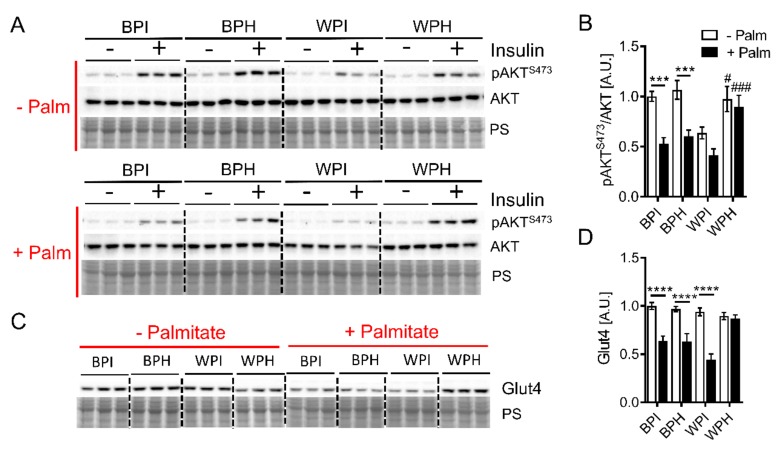
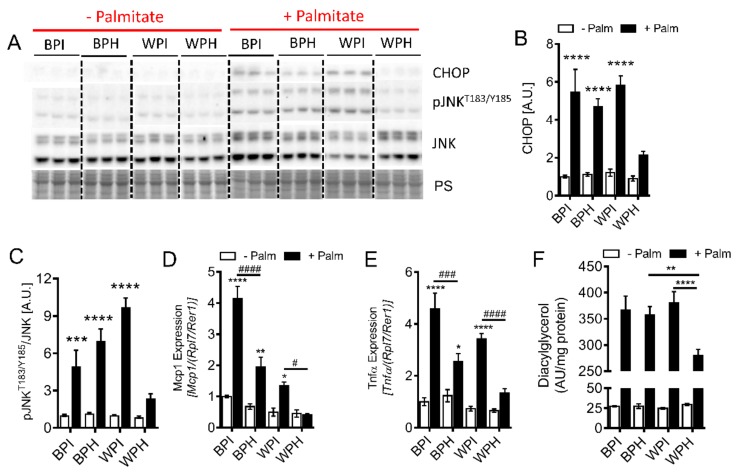
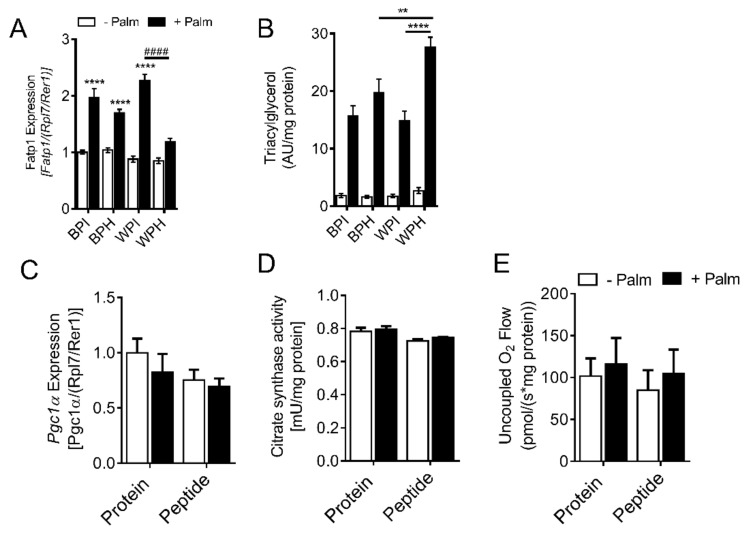
Deregulation of lipid metabolism and insulin function in muscle and adipose tissue are hallmarks of systemic insulin resistance, which can progress to type 2 diabetes. While previous studies suggested that milk proteins influence systemic glucose homeostasis and insulin function, it remains unclear whether bioactive peptides generated from whey alter lipid metabolism and its accumulation in muscle and adipose tissue. Therefore, we incubated murine 3T3-L1 preadipocytes and C2C12 myotubes with a whey peptide mixture produced through pepsin-pancreatin digestion, mimicking peptides generated in the gut from whey protein hydrolysis, and examined its effect on indicators of lipid metabolism and insulin sensitivity. Whey peptides, particularly those derived from bovine serum albumin (BSA), promoted 3T3-L1 adipocyte differentiation and triacylglycerol (TG) accumulation in accordance with peroxisome proliferator-activated receptor γ (PPARγ) upregulation. Whey/BSA peptides also increased lipolysis and mitochondrial fat oxidation in adipocytes, which was associated with the upregulation of peroxisome proliferator-activated receptor δ (PPARδ). In C2C12 myotubes, whey but not BSA peptides ameliorated palmitate-induced insulin resistance, which was associated with reduced inflammation and diacylglycerol accumulation, and increased sequestration of fatty acids in the TG pool. Taken together, our study suggests that whey peptides generated via pepsin-pancreatin digestion profoundly alter lipid metabolism and accumulation in adipocytes and skeletal myotubes.
Keywords: PPAR; adipocytes; differentiation; insulin resistance; lipolysis; lipotoxicity; metabolism; mitochondria; myocytes; whey peptides.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Griffin M.E., Marcucci M.J., Cline G.W., Bell K., Barucci N., Lee D., Goodyear L.J., Kraegen E.W., White M.F., Shulman G.I. Free fatty acid-induced insulin resistance is associated with activation of protein kinase C theta and alterations in the insulin signaling cascade. Diabetes. 1999;48:1270–1274. doi: 10.2337/diabetes.48.6.1270. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
