

ORP3 phosphorylation regulates phosphatidylinositol 4-phosphate and Ca2+ dynamics at plasma membrane-ER contact sites
- PMID: 32041906
- PMCID: PMC7097422
- DOI: 10.1242/jcs.237388
ORP3 phosphorylation regulates phosphatidylinositol 4-phosphate and Ca2+ dynamics at plasma membrane-ER contact sites
Abstract
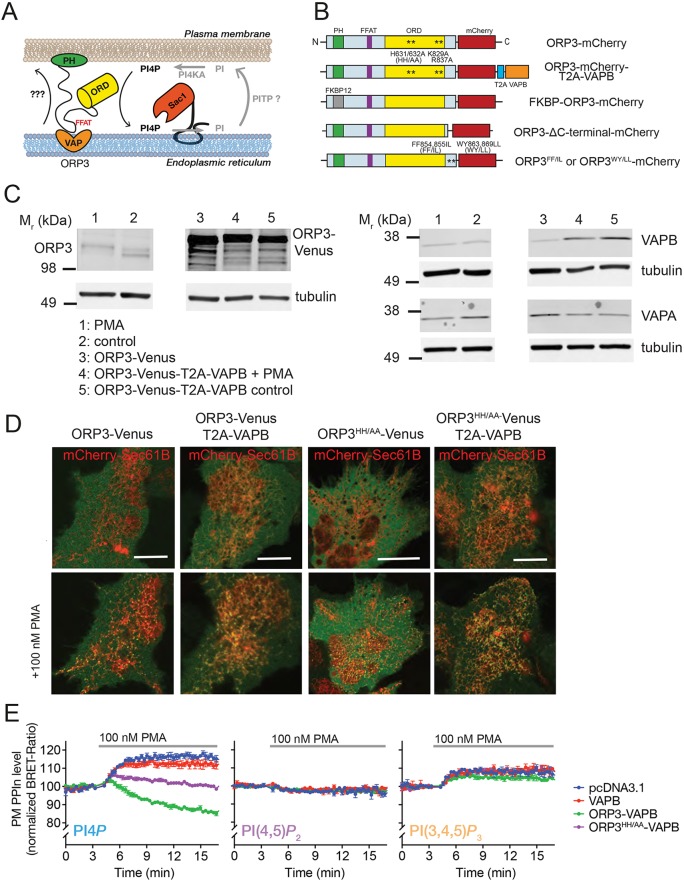
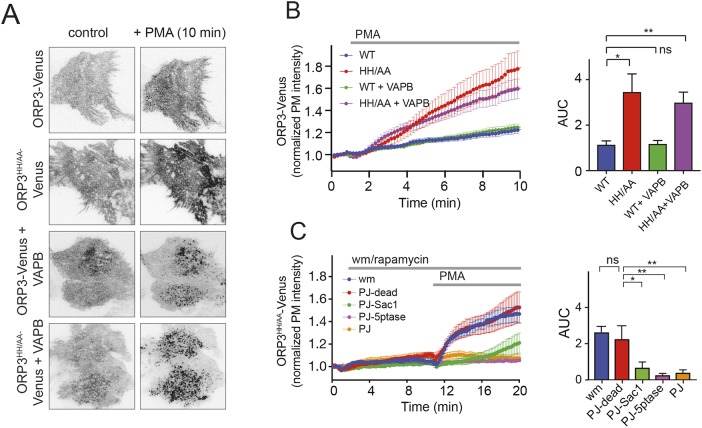
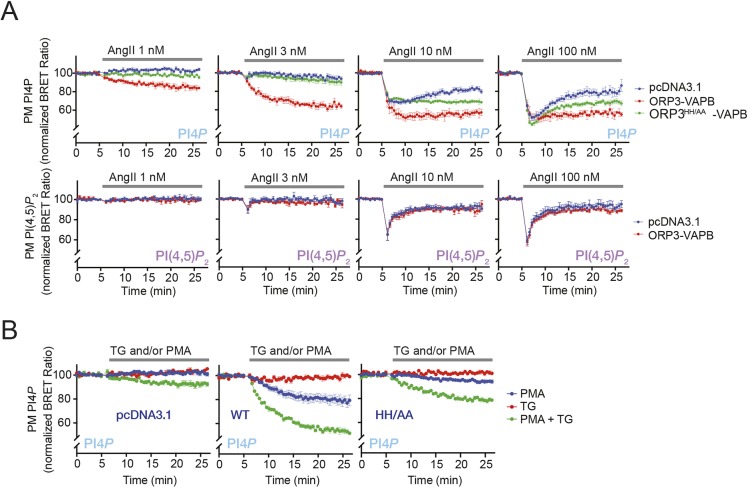
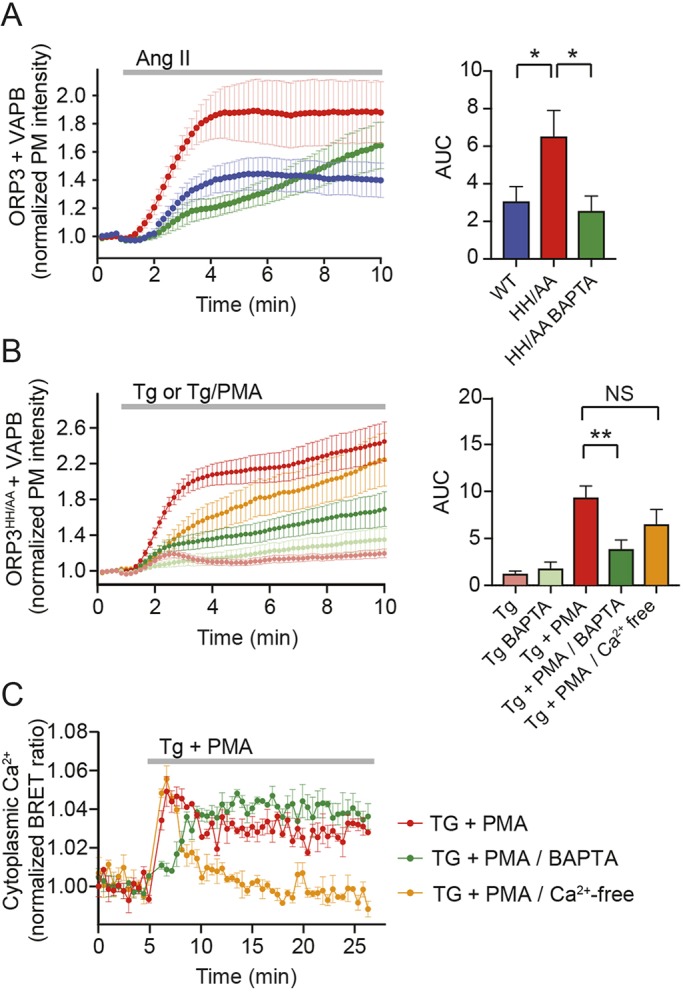
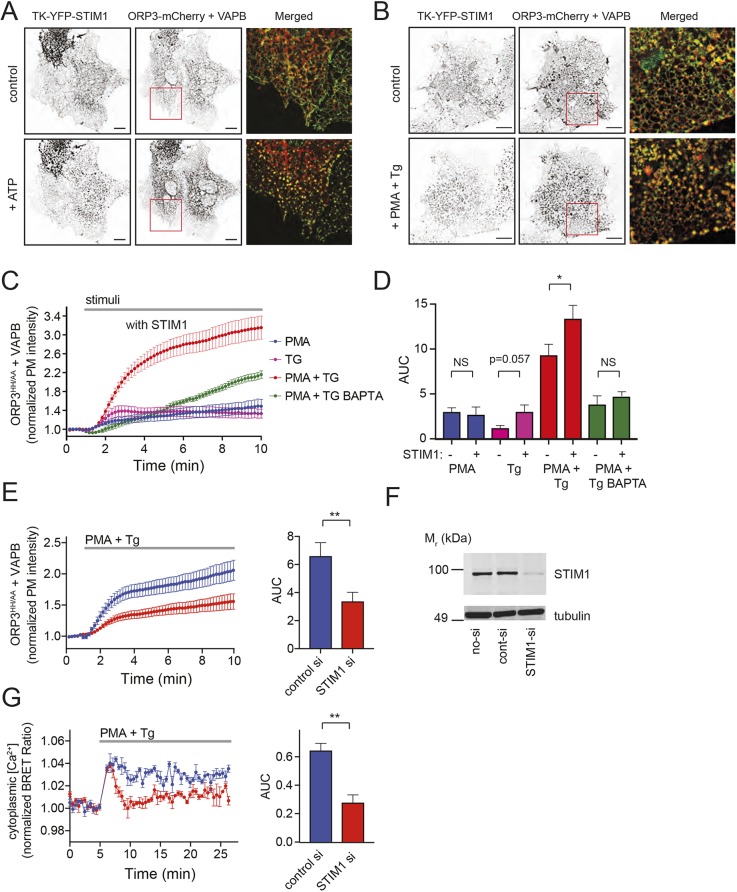
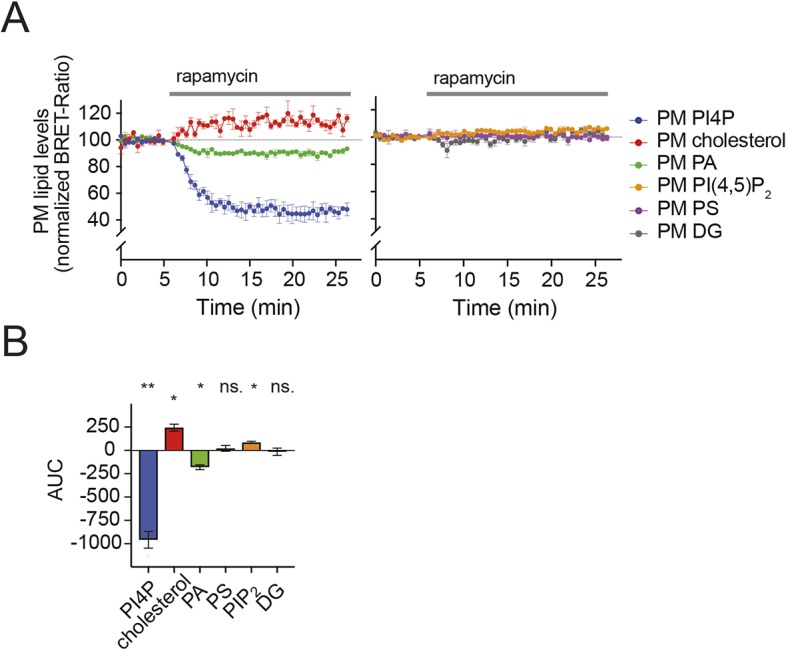
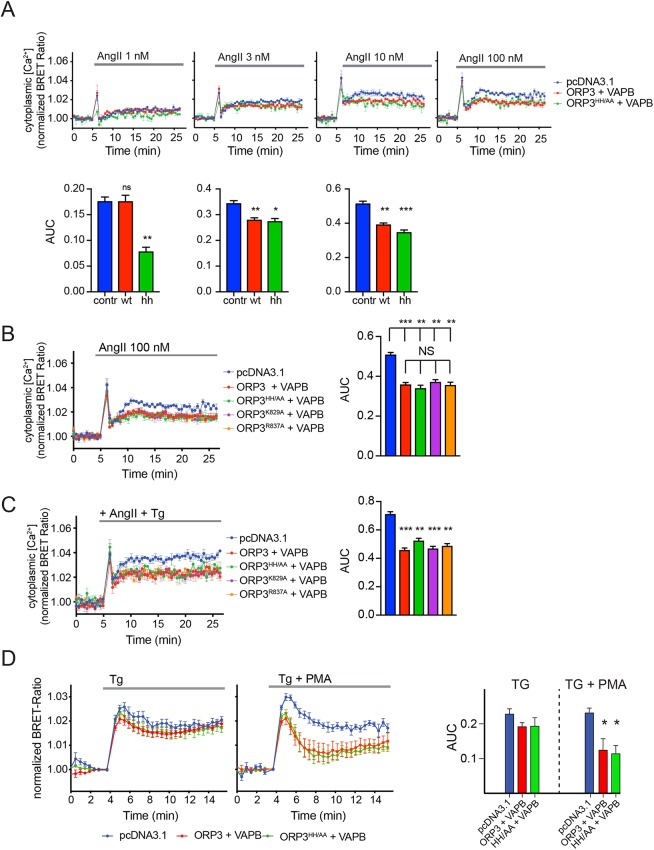
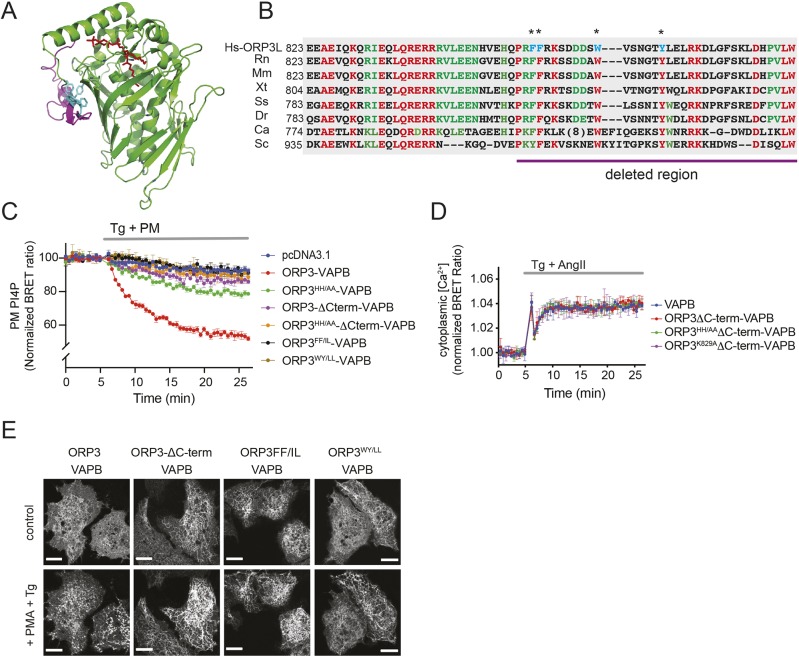
Oxysterol-binding protein (OSBP)-related proteins (ORPs) mediate non-vesicular lipid transfer between intracellular membranes. Phosphoinositide (PI) gradients play important roles in the ability of OSBP and some ORPs to transfer cholesterol and phosphatidylserine between the endoplasmic reticulum (ER) and other organelle membranes. Here, we show that plasma membrane (PM) association of ORP3 (also known as OSBPL3), a poorly characterized ORP family member, is triggered by protein kinase C (PKC) activation, especially when combined with Ca2+ increases, and is determined by both PI(4,5)P2 and PI4P After activation, ORP3 efficiently extracts PI4P and to a lesser extent phosphatidic acid from the PM, and slightly increases PM cholesterol levels. Full activation of ORP3 resulted in decreased PM PI4P levels and inhibited Ca2+ entry via the store-operated Ca2+ entry pathway. The C-terminal region of ORP3 that follows the strictly defined lipid transfer domain was found to be critical for the proper localization and function of the protein.
Keywords: BRET; ORP3; Phosphoinositides; Protein kinase C; SOCE; STIM1; TIRF.
© 2020. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
Oxysterol-binding protein-related protein (ORP) 6 localizes to the ER and ER-plasma membrane contact sites and is involved in the turnover of PI4P in cerebellar granule neurons.Exp Cell Res. 2018 Sep 15;370(2):601-612. doi: 10.1016/j.yexcr.2018.07.025. Epub 2018 Jul 18. Exp Cell Res. 2018. PMID: 30028970
-
LIPID transfer proteins regulate store-operated calcium entry via control of plasma membrane phosphoinositides.Cell Calcium. 2022 Sep;106:102631. doi: 10.1016/j.ceca.2022.102631. Epub 2022 Jul 11. Cell Calcium. 2022. PMID: 35853265 Free PMC article.
-
OSBP-Related Protein Family: Mediators of Lipid Transport and Signaling at Membrane Contact Sites.Int Rev Cell Mol Biol. 2016;321:299-340. doi: 10.1016/bs.ircmb.2015.09.006. Epub 2015 Nov 18. Int Rev Cell Mol Biol. 2016. PMID: 26811291 Review.
-
The crystal structure of ORP3 reveals the conservative PI4P binding pattern.Biochem Biophys Res Commun. 2020 Sep 3;529(4):1005-1010. doi: 10.1016/j.bbrc.2020.06.090. Epub 2020 Jul 30. Biochem Biophys Res Commun. 2020. PMID: 32819557
-
Ca2+ signaling and lipid transfer 'pas a deux' at ER-PM contact sites orchestrate cell migration.Cell Calcium. 2020 Jul;89:102226. doi: 10.1016/j.ceca.2020.102226. Epub 2020 Jun 1. Cell Calcium. 2020. PMID: 32505782 Review.
Cited by
-
Fatty Liver Disease-Alcoholic and Non-Alcoholic: Similar but Different.Int J Mol Sci. 2022 Dec 19;23(24):16226. doi: 10.3390/ijms232416226. Int J Mol Sci. 2022. PMID: 36555867 Free PMC article. Review.
-
Metabolic routing maintains the unique fatty acid composition of phosphoinositides.EMBO Rep. 2022 Jul 5;23(7):e54532. doi: 10.15252/embr.202154532. Epub 2022 Jun 16. EMBO Rep. 2022. PMID: 35712788 Free PMC article.
-
The Role of Lipids in CRAC Channel Function.Biomolecules. 2022 Feb 23;12(3):352. doi: 10.3390/biom12030352. Biomolecules. 2022. PMID: 35327543 Free PMC article. Review.
-
Differential activation of the inositol 5-phosphatase SHIP2 by EGF and insulin signaling pathways.J Biol Chem. 2025 Jul;301(7):110275. doi: 10.1016/j.jbc.2025.110275. Epub 2025 May 22. J Biol Chem. 2025. PMID: 40412518 Free PMC article.
-
Endoplasmic Reticulum-Plasma Membrane Contact Sites as an Organizing Principle for Compartmentalized Calcium and cAMP Signaling.Int J Mol Sci. 2021 Apr 29;22(9):4703. doi: 10.3390/ijms22094703. Int J Mol Sci. 2021. PMID: 33946838 Free PMC article. Review.
References
-
- Bojjireddy N., Botyanszki J., Hammond G., Creech D., Peterson R., Kemp D. C., Snead M., Brown R., Morrison A., Wilson S. et al. (2014). Pharmacological and genetic targeting of the PI4KA enzyme reveals its important role in maintaining plasma membrane phosphatidylinositol 4-phosphate and phosphatidylinositol 4,5-bisphosphate levels. J. Biol. Chem. 289, 6120-6132. 10.1074/jbc.M113.531426 - DOI - PMC - PubMed
-
- Broad L. M., Braun F.-J., Lievremont J.-P., Bird G. S. J., Kurosaki T. and Putney J. W. Jr (2001). Role of the phospholipase C-inositol 1,4,5-trisphosphate pathway in calcium release-activated calcium current and capacitative calcium entry. J. Biol. Chem. 276, 15945-15952. 10.1074/jbc.M011571200 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
