

Nanocatalytic activity of clean-surfaced, faceted nanocrystalline gold enhances remyelination in animal models of multiple sclerosis
- PMID: 32041968
- PMCID: PMC7010780
- DOI: 10.1038/s41598-020-58709-w
Nanocatalytic activity of clean-surfaced, faceted nanocrystalline gold enhances remyelination in animal models of multiple sclerosis
Abstract
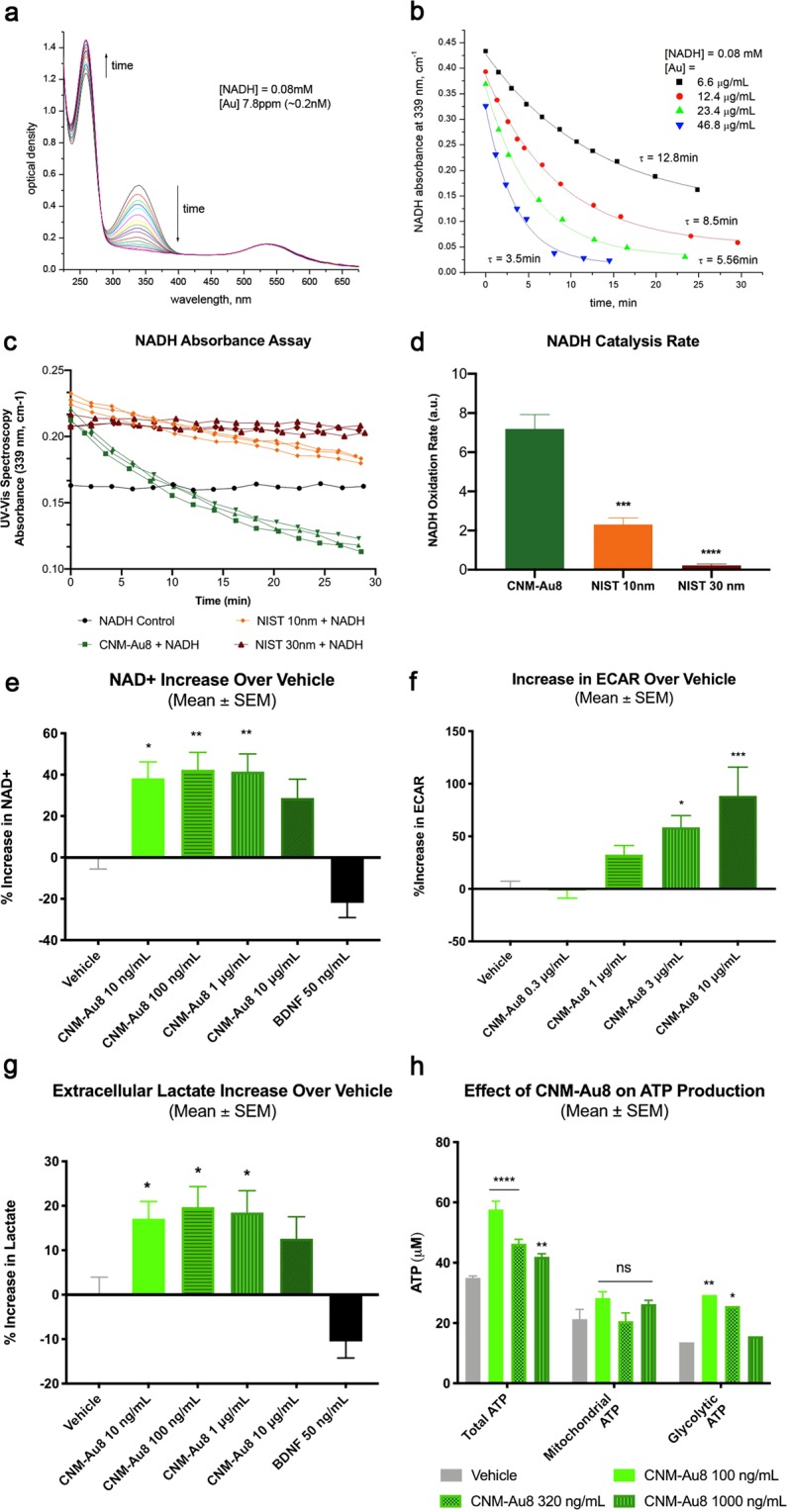
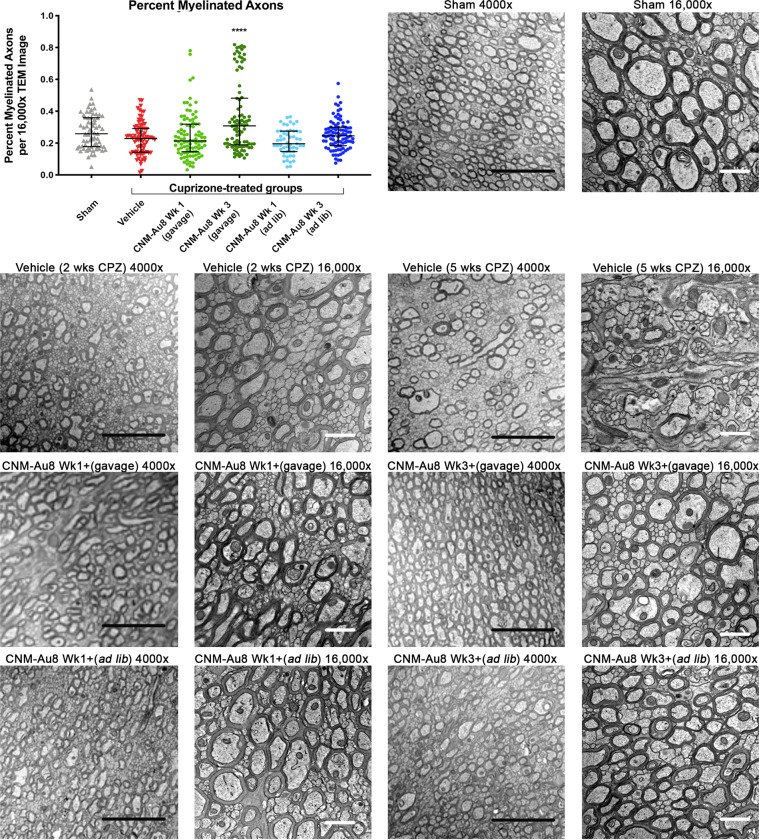
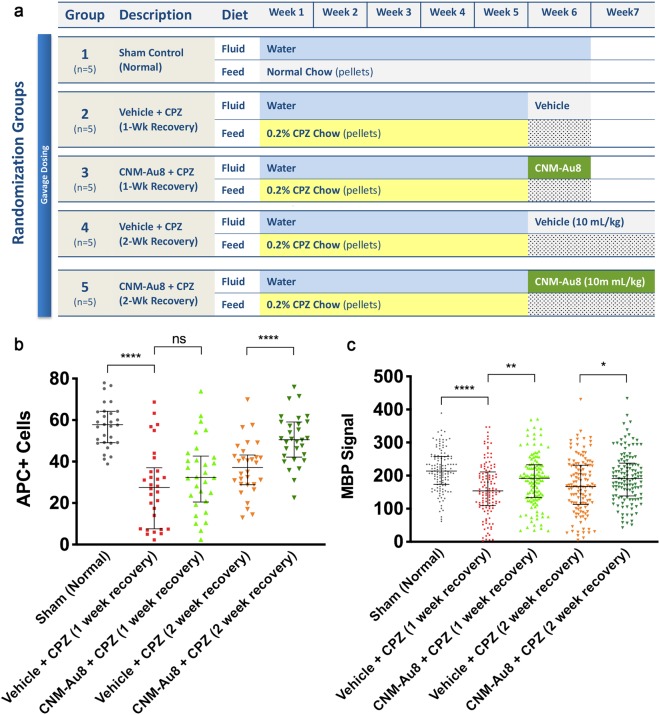
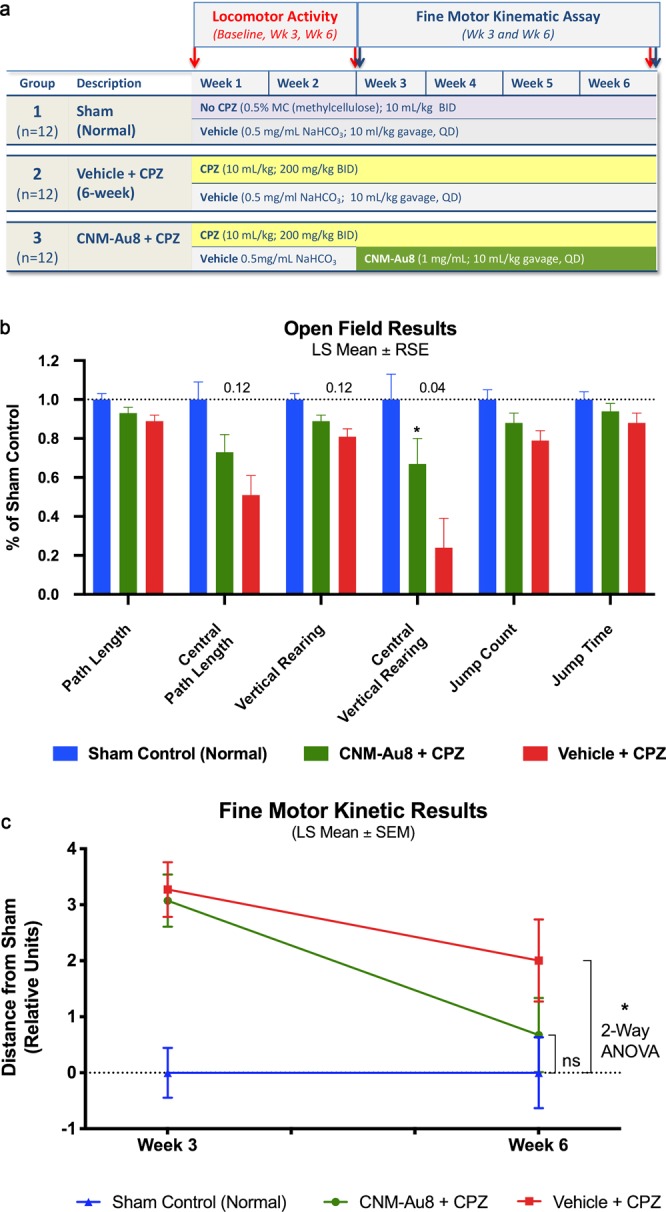
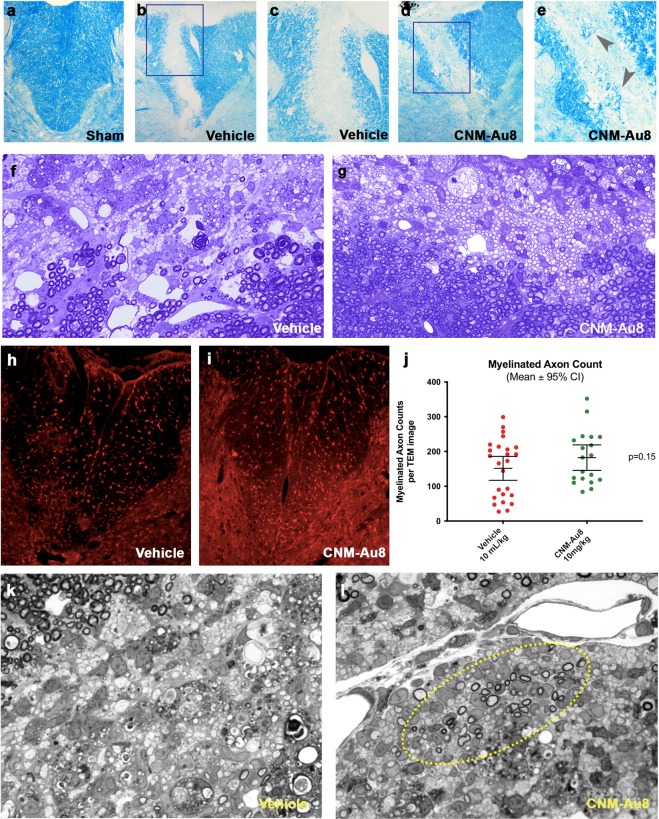
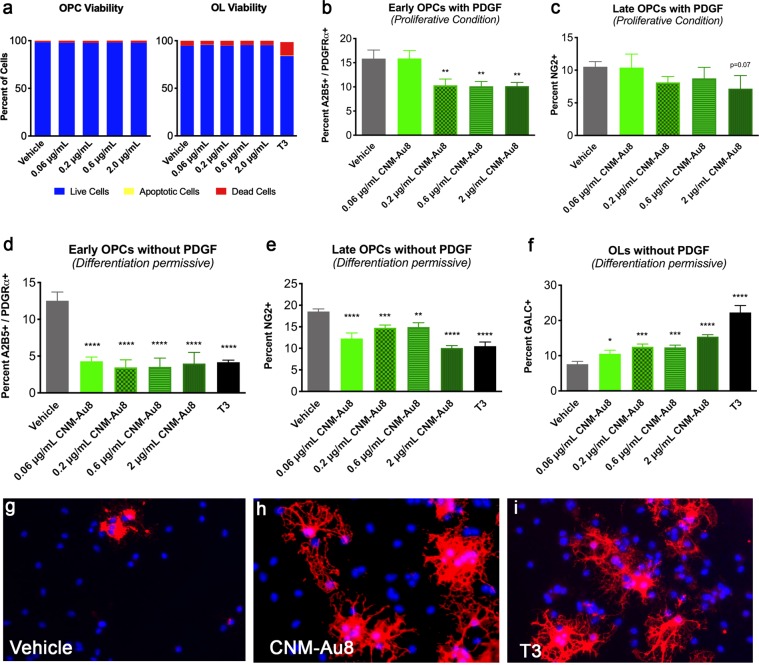
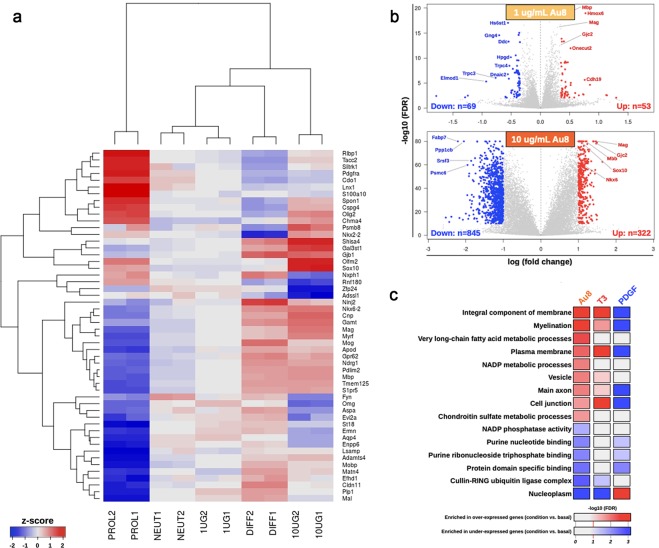
Development of pharmacotherapies that promote remyelination is a high priority for multiple sclerosis (MS), due to their potential for neuroprotection and restoration of function through repair of demyelinated lesions. A novel preparation of clean-surfaced, faceted gold nanocrystals demonstrated robust remyelinating activity in response to demyelinating agents in both chronic cuprizone and acute lysolecithin rodent animal models. Furthermore, oral delivery of gold nanocrystals improved motor functions of cuprizone-treated mice in both open field and kinematic gait studies. Gold nanocrystal treatment of oligodendrocyte precursor cells in culture resulted in oligodendrocyte maturation and expression of myelin differentiation markers. Additional in vitro data demonstrated that these gold nanocrystals act via a novel energy metabolism pathway involving the enhancement of key indicators of aerobic glycolysis. In response to gold nanocrystals, co-cultured central nervous system cells exhibited elevated levels of the redox coenzyme nicotine adenine dinucleotide (NAD+), elevated total intracellular ATP levels, and elevated extracellular lactate levels, along with upregulation of myelin-synthesis related genes, collectively resulting in functional myelin generation. Based on these preclinical studies, clean-surfaced, faceted gold nanocrystals represent a novel remyelinating therapeutic for multiple sclerosis.
Conflict of interest statement
J.Z.Z., M.M., A.R.D., K.S.H., M.T.H. and M.G.M. are employees of Clene Nanomedicine and receive salary and stock options. All other authors have no competing interests to declare.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
