

Development of a covalent inhibitor of gut bacterial bile salt hydrolases
- PMID: 32042200
- PMCID: PMC7036035
- DOI: 10.1038/s41589-020-0467-3
Development of a covalent inhibitor of gut bacterial bile salt hydrolases
Abstract
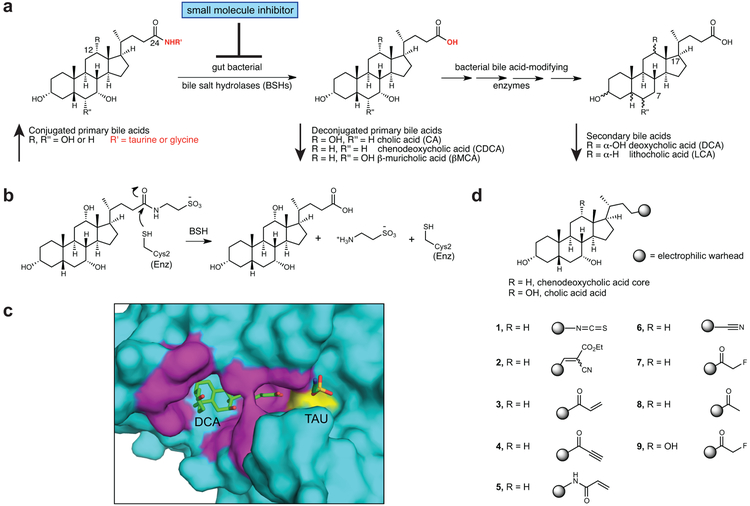
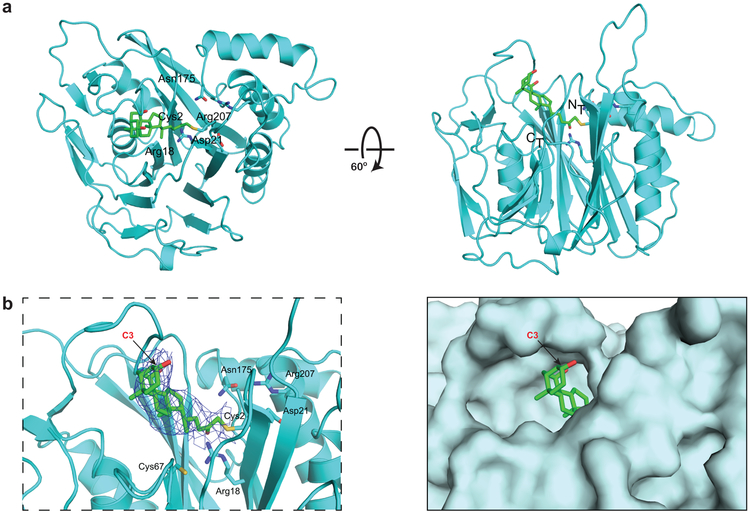
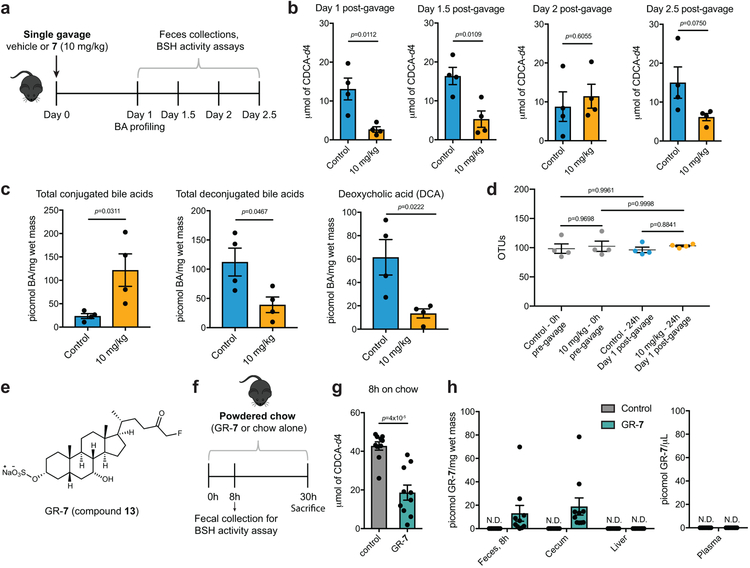
Bile salt hydrolase (BSH) enzymes are widely expressed by human gut bacteria and catalyze the gateway reaction leading to secondary bile acid formation. Bile acids regulate key metabolic and immune processes by binding to host receptors. There is an unmet need for a potent tool to inhibit BSHs across all gut bacteria to study the effects of bile acids on host physiology. Here, we report the development of a covalent pan-inhibitor of gut bacterial BSHs. From a rationally designed candidate library, we identified a lead compound bearing an alpha-fluoromethyl ketone warhead that modifies BSH at the catalytic cysteine residue. This inhibitor abolished BSH activity in conventional mouse feces. Mice gavaged with a single dose of this compound displayed decreased BSH activity and decreased deconjugated bile acid levels in feces. Our studies demonstrate the potential of a covalent BSH inhibitor to modulate bile acid composition in vivo.
Figures


Similar articles
-
A Gut-Restricted Lithocholic Acid Analog as an Inhibitor of Gut Bacterial Bile Salt Hydrolases.ACS Chem Biol. 2021 Aug 20;16(8):1401-1412. doi: 10.1021/acschembio.1c00192. Epub 2021 Jul 19. ACS Chem Biol. 2021. PMID: 34279901 Free PMC article.
-
Metagenomic analysis of the human microbiome reveals the association between the abundance of gut bile salt hydrolases and host health.Gut Microbes. 2020 Sep 2;11(5):1300-1313. doi: 10.1080/19490976.2020.1748261. Epub 2020 Apr 24. Gut Microbes. 2020. PMID: 32329665 Free PMC article.
-
Taxonomic profiling and populational patterns of bacterial bile salt hydrolase (BSH) genes based on worldwide human gut microbiome.Microbiome. 2019 Jan 23;7(1):9. doi: 10.1186/s40168-019-0628-3. Microbiome. 2019. PMID: 30674356 Free PMC article.
-
New insights into bacterial bile resistance mechanisms: the role of bile salt hydrolase and its impact on human health.Food Res Int. 2018 Oct;112:250-262. doi: 10.1016/j.foodres.2018.06.035. Epub 2018 Jun 20. Food Res Int. 2018. PMID: 30131136 Review.
-
Interactions between gut bacteria and bile in health and disease.Mol Aspects Med. 2017 Aug;56:54-65. doi: 10.1016/j.mam.2017.06.002. Epub 2017 Jun 21. Mol Aspects Med. 2017. PMID: 28602676 Review.
Cited by
-
Thalidomide mitigates Crohn's disease colitis by modulating gut microbiota, metabolites, and regulatory T cell immunity.J Pharm Anal. 2025 Apr;15(4):101121. doi: 10.1016/j.jpha.2024.101121. Epub 2024 Oct 18. J Pharm Anal. 2025. PMID: 40309194 Free PMC article.
-
Microbiota mechanisms in cancer progression and therapy.Cell Chem Biol. 2025 May 15;32(5):653-677. doi: 10.1016/j.chembiol.2025.04.005. Epub 2025 May 6. Cell Chem Biol. 2025. PMID: 40334660 Review.
-
The changing metabolic landscape of bile acids - keys to metabolism and immune regulation.Nat Rev Gastroenterol Hepatol. 2024 Jul;21(7):493-516. doi: 10.1038/s41575-024-00914-3. Epub 2024 Apr 4. Nat Rev Gastroenterol Hepatol. 2024. PMID: 38575682 Free PMC article. Review.
-
Chemoproteomic Approaches for Unraveling Prokaryotic Biology.Isr J Chem. 2023 Mar;63(3-4):e202200076. doi: 10.1002/ijch.202200076. Epub 2023 Feb 28. Isr J Chem. 2023. PMID: 37842282 Free PMC article.
-
Defective humoral immunity disrupts bile acid homeostasis which promotes inflammatory disease of the small bowel.Nat Commun. 2022 Jan 26;13(1):525. doi: 10.1038/s41467-022-28126-w. Nat Commun. 2022. PMID: 35082296 Free PMC article.
References
-
- Ridlon JM, Kang D-J & Hylemon PB Bile salt biotransformations by human intestinal bacteria. J. Lipid Res 47, 241–259 (2006). - PubMed
-
- Fiorucci S & Distrutti E Bile Acid-Activated Receptors, Intestinal Microbiota, and the Treatment of Metabolic Disorders. Trends Mol Med 21, 702–714 (2015). - PubMed
-
- Setchell KD, Lawson AM, Tanida N & Sjövall J General methods for the analysis of metabolic profiles of bile acids and related compounds in feces. J. Lipid Res 24, 1085–1100 (1983). - PubMed
-
- Hamilton JP et al. Human cecal bile acids: concentration and spectrum. Am. J. Physiol. Gastrointest. Liver Physiol 293, G256–G263 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
