

Computational saturation mutagenesis to predict structural consequences of systematic mutations in the beta subunit of RNA polymerase in Mycobacterium leprae
- PMID: 32042379
- PMCID: PMC7000446
- DOI: 10.1016/j.csbj.2020.01.002
Computational saturation mutagenesis to predict structural consequences of systematic mutations in the beta subunit of RNA polymerase in Mycobacterium leprae
Abstract
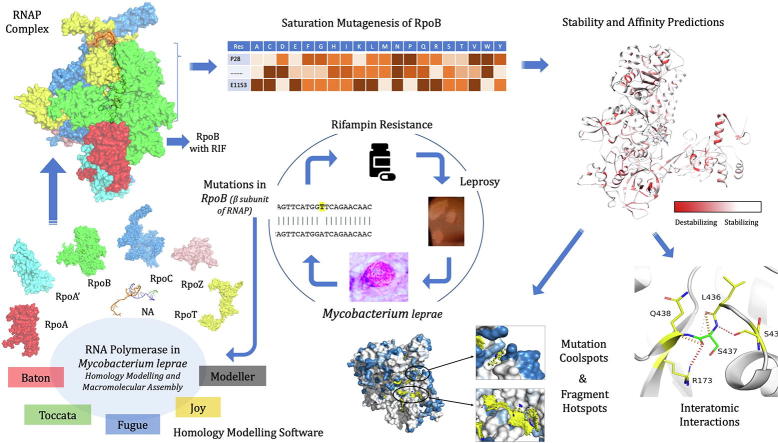
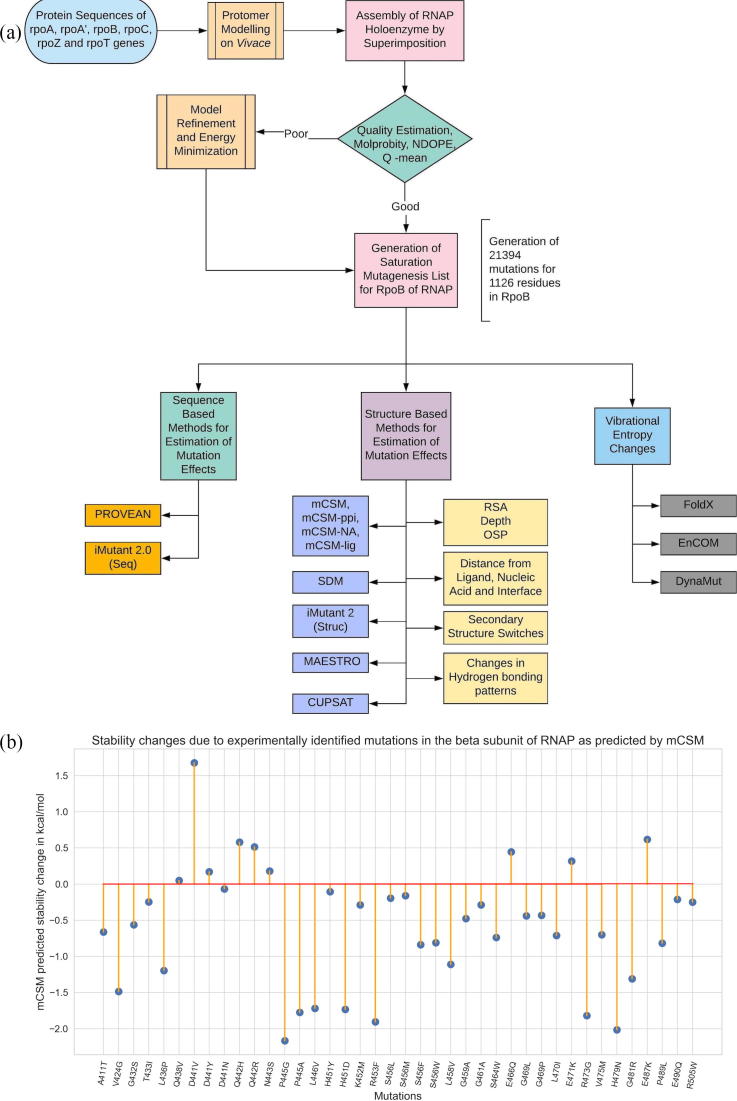
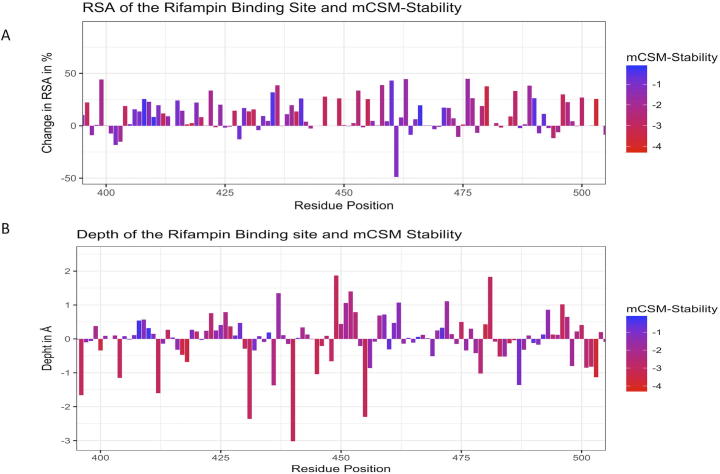
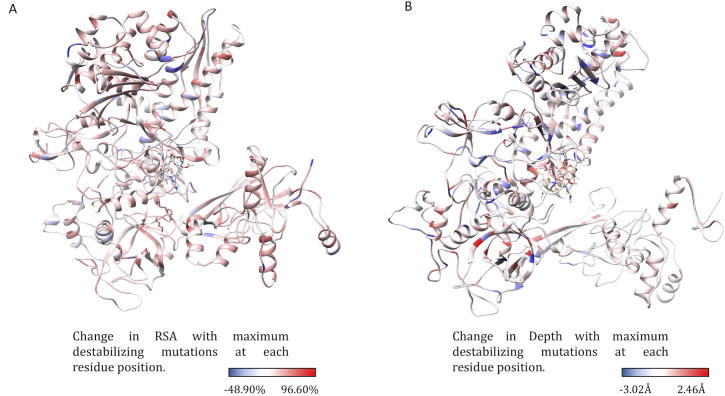
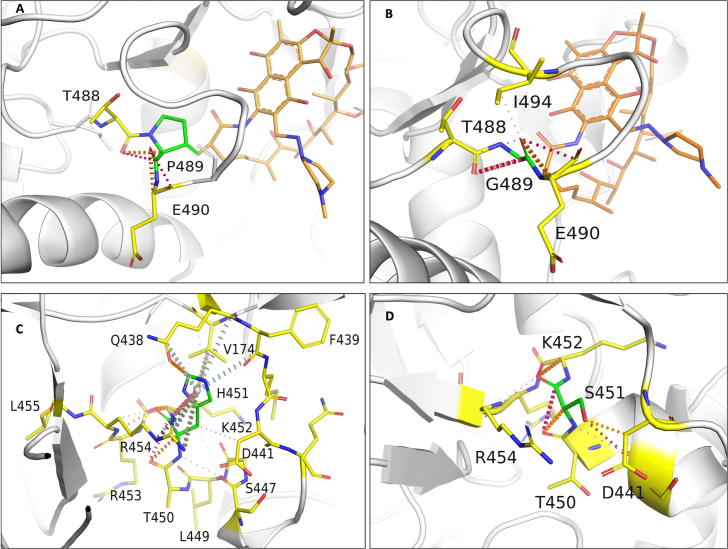
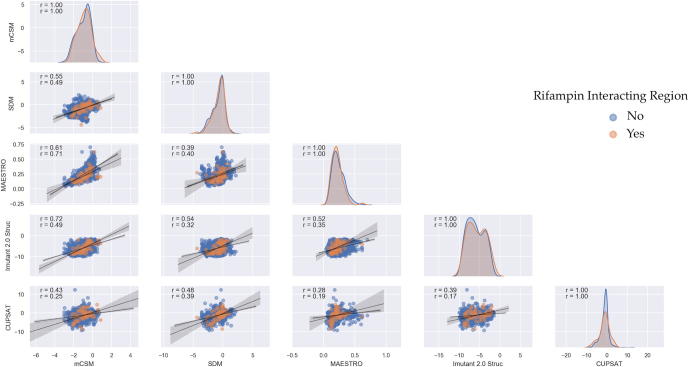
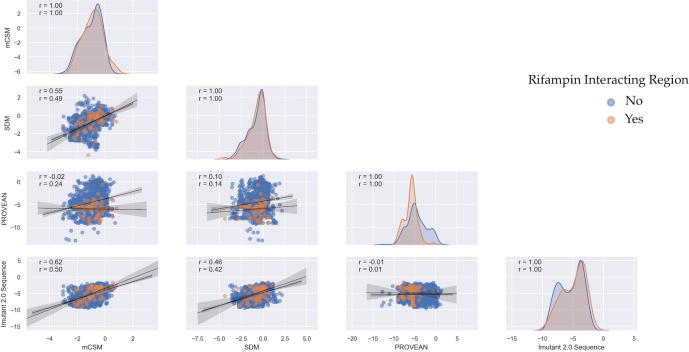
Rifampin resistance in leprosy may remain undetected due to the lack of rapid and effective diagnostic tools. A quick and reliable method is essential to determine the impacts of emerging detrimental mutations in the drug targets. The functional consequences of missense mutations in the β-subunit of RNA polymerase (RNAP) in Mycobacterium leprae (M. leprae) contribute to phenotypic resistance to rifampin in leprosy. Here, we report in-silico saturation mutagenesis of all residues in the β-subunit of RNAP to all other 19 amino acid types (generating 21,394 mutations for 1126 residues) and predict their impacts on overall thermodynamic stability, on interactions at subunit interfaces, and on β-subunit-RNA and rifampin affinities (only for the rifampin binding site) using state-of-the-art structure, sequence and normal mode analysis-based methods. Mutations in the conserved residues that line the active-site cleft show largely destabilizing effects, resulting in increased relative solvent accessibility and a concomitant decrease in residue-depth (the extent to which a residue is buried in the protein structure space) of the mutant residues. The mutations at residue positions S437, G459, H451, P489, K884 and H1035 are identified as extremely detrimental as they induce highly destabilizing effects on the overall protein stability, and nucleic acid and rifampin affinities. Destabilizing effects were predicted for all the clinically/experimentally identified rifampin-resistant mutations in M. leprae indicating that this model can be used as a surveillance tool to monitor emerging detrimental mutations that destabilise RNAP-rifampin interactions and confer rifampin resistance in leprosy.
Author summary: The emergence of primary and secondary drug resistance to rifampin in leprosy is a growing concern and poses a threat to the leprosy control and elimination measures globally. In the absence of an effective in-vitro system to detect and monitor phenotypic resistance to rifampin in leprosy, diagnosis mainly relies on the presence of mutations in drug resistance determining regions of the rpoB gene that encodes the β-subunit of RNAP in M. leprae. Few labs in the world perform mouse food pad propagation of M. leprae in the presence of drugs (rifampin) to determine growth patterns and confirm resistance, however the duration of these methods lasts from 8 to 12 months making them impractical for diagnosis. Understanding molecular mechanisms of drug resistance is vital to associating mutations to clinically detected drug resistance in leprosy. Here we propose an in-silico saturation mutagenesis approach to comprehensively elucidate the structural implications of any mutations that exist or that can arise in the β-subunit of RNAP in M. leprae. Most of the predicted mutations may not occur in M. leprae due to fitness costs but the information thus generated by this approach help decipher the impacts of mutations across the structure and conversely enable identification of stable regions in the protein that are least impacted by mutations (mutation coolspots) which can be a potential choice for small molecule binding and structure guided drug discovery.
Keywords: In-silico Saturation Mutagenesis; Mutation Coolspots; Mycobacterium leprae; RNA Polymerase; Rifampin; Thermodynamic stability.
© 2020 The Authors.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures

References
-
- World Health Organization. Global tuberculosis report 2018 [Internet]. World Health Organization; 2018 [cited 2018 Dec 19]. 231 p. Available from: http://apps.who.int/iris/handle/10665/274453
-
- WHO | Weekly Epidemiological Record, 31 August 2018, vol. 93, 35 (pp. 444–456) [Internet]. WHO. [cited 2018 Dec 19]. Available from: http://www.who.int/wer/2018/wer9335/en/.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources