

Postural control of arm and fingers through integration of movement commands
- PMID: 32043973
- PMCID: PMC7062460
- DOI: 10.7554/eLife.52507
Postural control of arm and fingers through integration of movement commands
Abstract
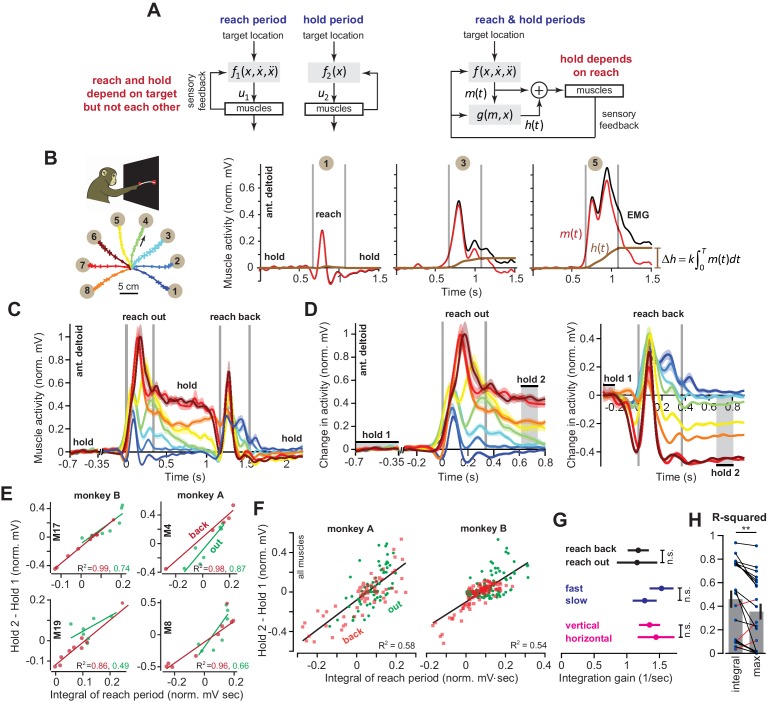
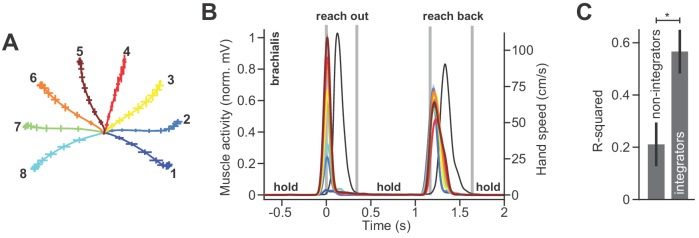
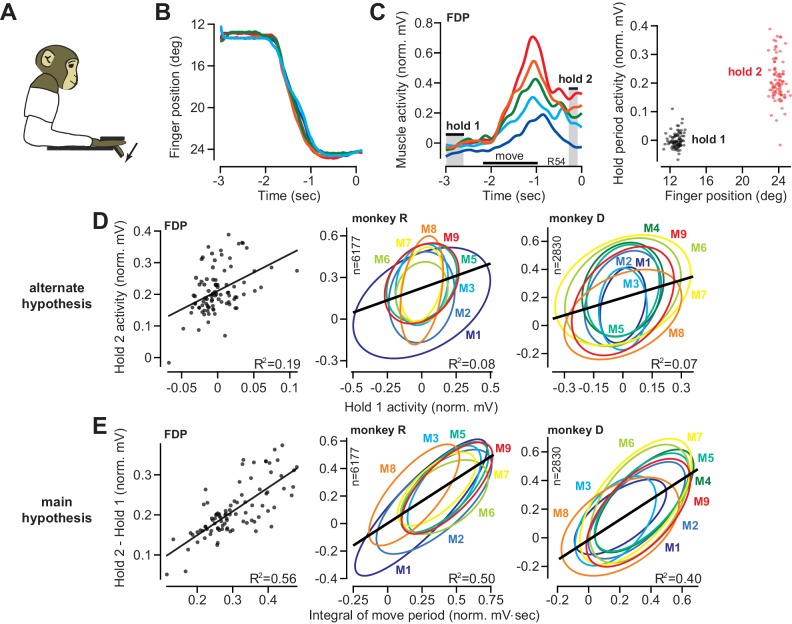
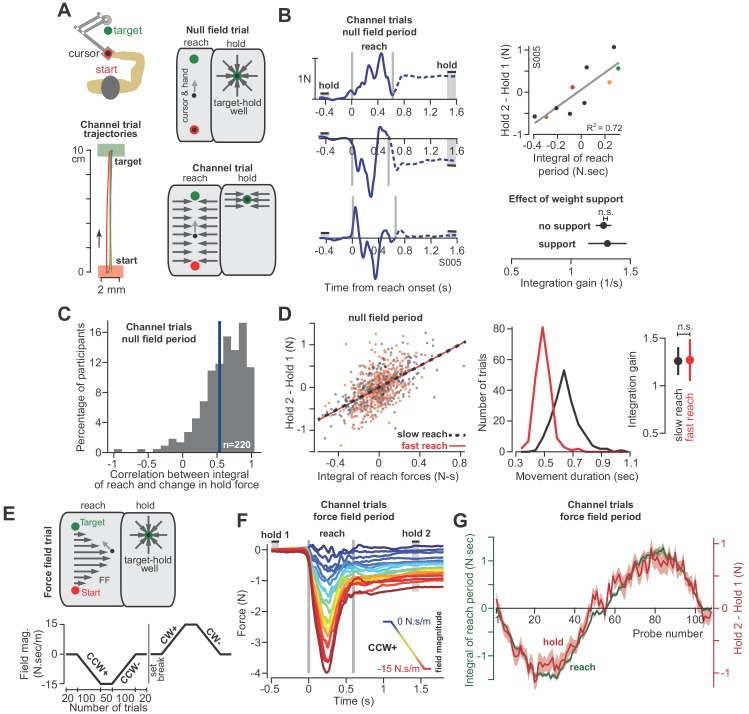
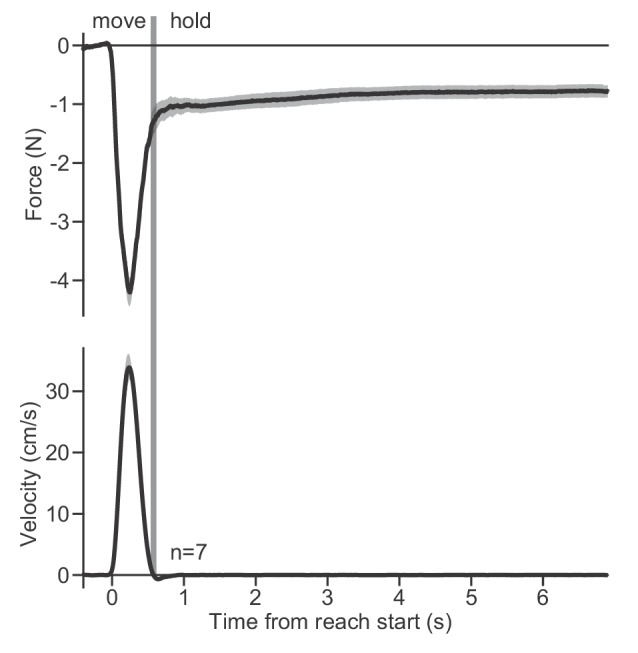
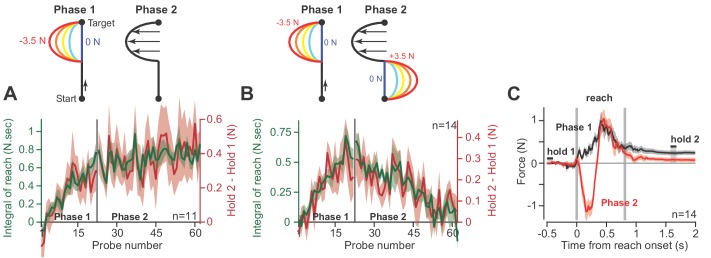
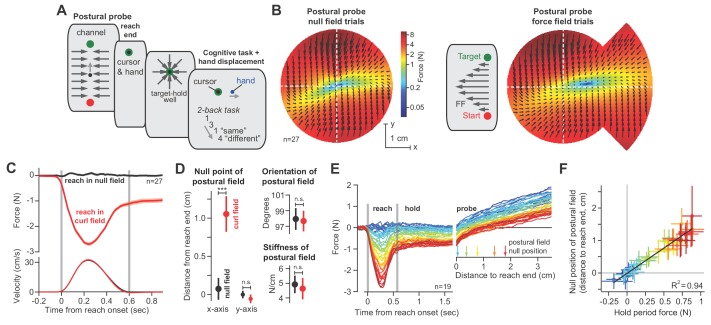
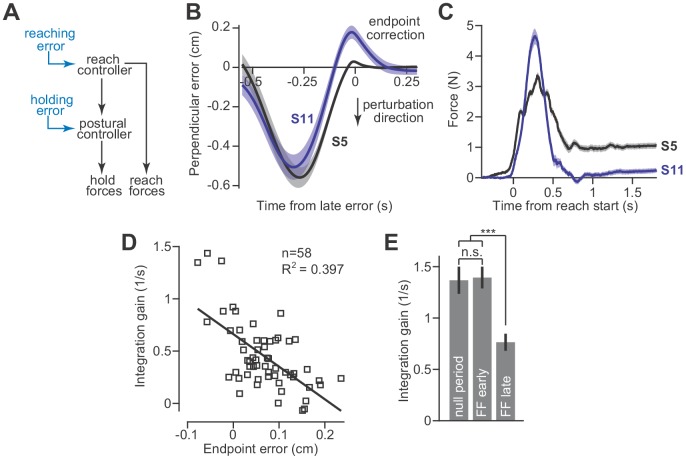
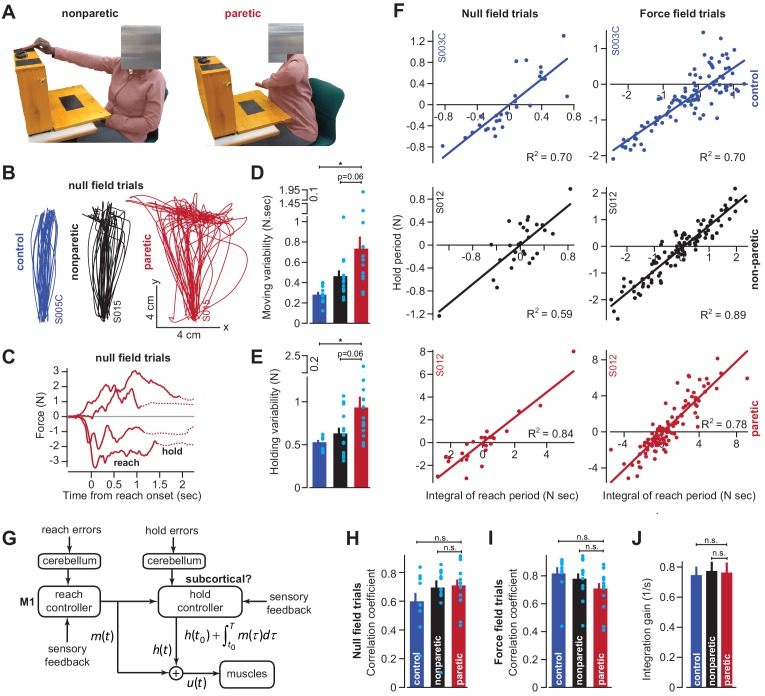
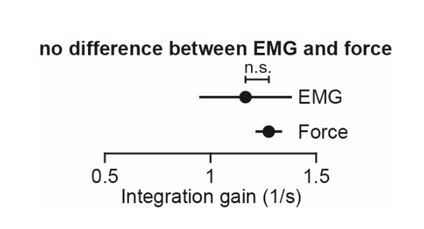
Every movement ends in a period of stillness. Current models assume that commands that hold the limb at a target location do not depend on the commands that moved the limb to that location. Here, we report a surprising relationship between movement and posture in primates: on a within-trial basis, the commands that hold the arm and finger at a target location depend on the mathematical integration of the commands that moved the limb to that location. Following damage to the corticospinal tract, both the move and hold period commands become more variable. However, the hold period commands retain their dependence on the integral of the move period commands. Thus, our data suggest that the postural controller possesses a feedforward module that uses move commands to calculate a component of hold commands. This computation may arise within an unknown subcortical system that integrates cortical commands to stabilize limb posture.
Keywords: Reaching; human; integration; motor control; motor learning; neuroscience; posture; rhesus macaque; stroke.
Plain language summary
Moving an arm requires the brain to send electrical signals to the arm’s muscles, causing them to contract. Neuroscientists call these types of brain signals “move signals”. The brain also sends so-called hold signals, which hold the arm still in a desired position. Part of the brain known as the primary motor cortex helps to calculate the move signals for the arm, but it was unclear how the brain produces the corresponding hold signals. Fortunately, the fact that the brain moves other things besides arms may help answer this question. Previous research has shown, for example, that a brain area called the “neural integrator” calculates the hold signals needed to hold the eye in a specific position. The neural integrator does this by using basic principles of physics, and details of the speed and duration of the eye’s movements. Now, Albert et al. show a similar mechanism appears to control hold signals for arm movements. In one set of experiments, muscle activity was measured as monkeys moved their arms or fingers to different target positions. In other experiments, human volunteers held a robot arm, and Albert et al. measured the forces they produced while reaching and holding still. Both the human and monkey experiments revealed a relationship between move signals and hold signals. Like for eye movements, hold signals for the arm could be calculated from the move signals. In further experiments with stroke patients where the brain had been damaged, the move signals were found to be deteriorated, but the way hold signals were calculated stayed the same. This suggests that there is an unknown structure within the brain that calculates hold signals based on move signals. Investigating how the brain holds the arm still may help scientists understand why some neurological conditions like stroke or dystonia cause unwanted movements or unusual postures. This might also lead scientists to develop new ways to treat these conditions.
© 2020, Albert et al.
Conflict of interest statement
SA, AH, JJ, AZ, DS, SB, MC, JK, RS No competing interests declared
Figures
References
-
- Coderre AM, Zeid AA, Dukelow SP, Demmer MJ, Moore KD, Demers MJ, Bretzke H, Herter TM, Glasgow JI, Norman KE, Bagg SD, Scott SH. Assessment of upper-limb sensorimotor function of subacute stroke patients using visually guided reaching. Neurorehabilitation and Neural Repair. 2010;24:528–541. doi: 10.1177/1545968309356091. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
- R01NS078311/NS/NINDS NIH HHS/United States
- 1U19NS104649/NS/NINDS NIH HHS/United States
- F31 NS095706/NS/NINDS NIH HHS/United States
- U19 NS104649/NS/NINDS NIH HHS/United States
- F32 NS092350/NS/NINDS NIH HHS/United States
- R01 NS100066/NS/NINDS NIH HHS/United States
- F31NS095706/NS/NINDS NIH HHS/United States
- P30 EY019007/EY/NEI NIH HHS/United States
- MR/K023012/1/MRC_/Medical Research Council/United Kingdom
- MR/P023967/1/MRC_/Medical Research Council/United Kingdom
- F32NS092350/NS/NINDS NIH HHS/United States
- 1DP2NS083037/NS/NINDS NIH HHS/United States
- DP2 NS083037/NS/NINDS NIH HHS/United States
- MR/P023967/MRC_/Medical Research Council/United Kingdom
- R01NS100066/NS/NINDS NIH HHS/United States
- R01 NS078311/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
