

Characterizing lineage-specific evolution and the processes driving genomic diversification in chordates
- PMID: 32046633
- PMCID: PMC7011509
- DOI: 10.1186/s12862-020-1585-y
Characterizing lineage-specific evolution and the processes driving genomic diversification in chordates
Abstract
Background: Understanding the origins of genome content has long been a goal of molecular evolution and comparative genomics. By examining genome evolution through the guise of lineage-specific evolution, it is possible to make inferences about the evolutionary events that have given rise to species-specific diversification. Here we characterize the evolutionary trends found in chordate species using The Adaptive Evolution Database (TAED). TAED is a database of phylogenetically indexed gene families designed to detect episodes of directional or diversifying selection across chordates. Gene families within the database have been assessed for lineage-specific estimates of dN/dS and have been reconciled to the chordate species to identify retained duplicates. Gene families have also been mapped to the functional pathways and amino acid changes which occurred on high dN/dS lineages have been mapped to protein structures.
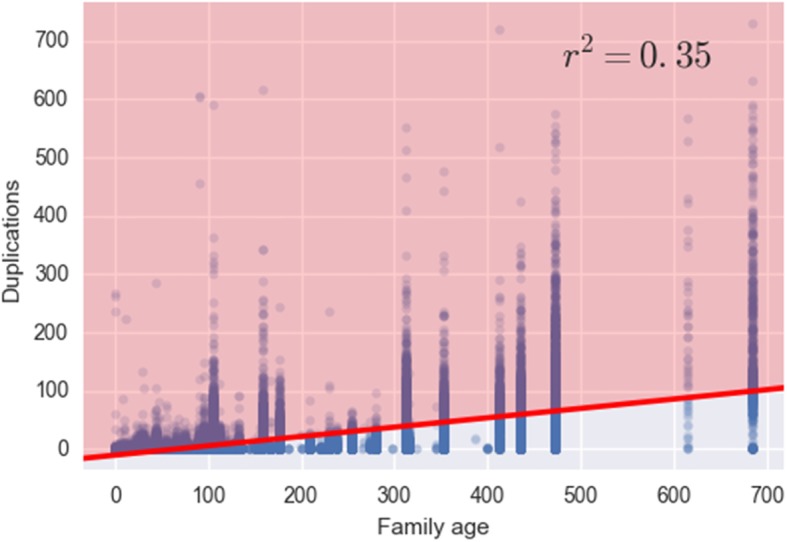
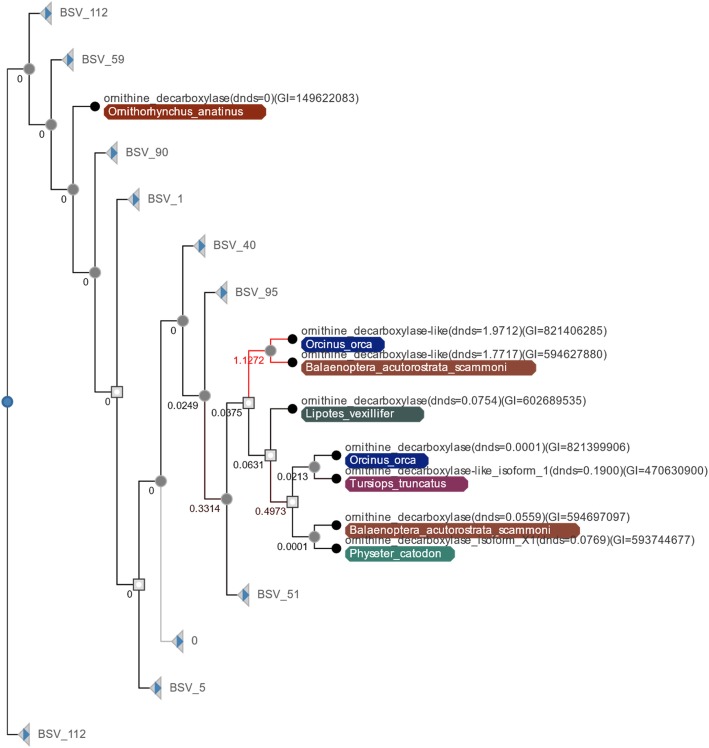
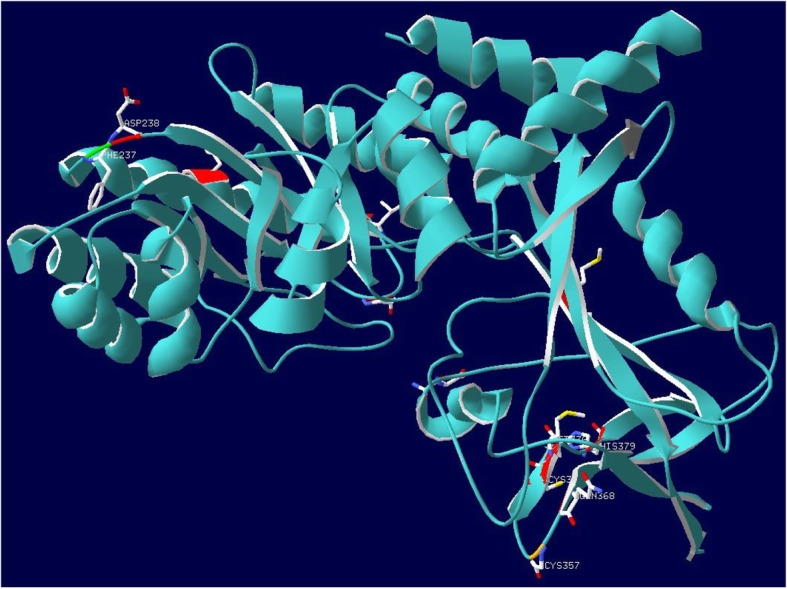
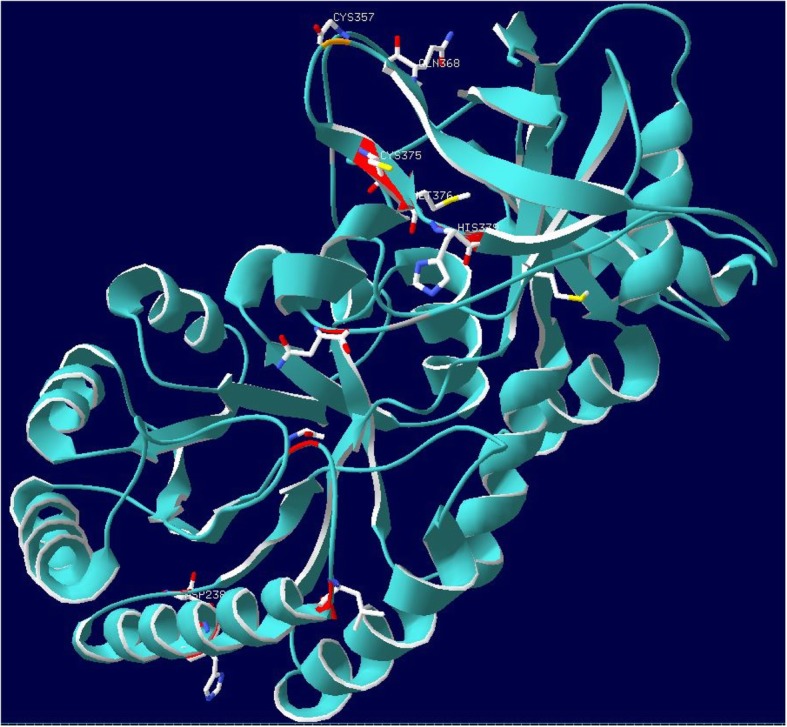
Results: An analysis of this exhaustive database has enabled a characterization of the processes of lineage-specific diversification in chordates. A pathway level enrichment analysis of TAED determined that pathways most commonly found to have elevated rates of evolution included those involved in metabolism, immunity, and cell signaling. An analysis of protein fold presence on proteins, after normalizing for frequency in the database, found common folds such as Rossmann folds, Jelly Roll folds, and TIM barrels were overrepresented on proteins most likely to undergo directional selection. A set of gene families which experience increased numbers of duplications within short evolutionary times are associated with pathways involved in metabolism, olfactory reception, and signaling. An analysis of protein secondary structure indicated more relaxed constraint in β-sheets and stronger constraint on alpha Helices, amidst a general preference for substitutions at exposed sites. Lastly a detailed analysis of the ornithine decarboxylase gene family, a key enzyme in the pathway for polyamine synthesis, revealed lineage-specific evolution along the lineage leading to Cetacea through rapid sequence evolution in a duplicate gene with amino acid substitutions causing active site rearrangement.
Conclusion: Episodes of lineage-specific evolution are frequent throughout chordate species. Both duplication and directional selection have played large roles in the evolution of the phylum. TAED is a powerful tool for facilitating this understanding of lineage-specific evolution.
Keywords: Comparative genomics; Gene duplication; Molecular evolution; Pathway evolution; Protein structure.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Altenhoff AM, Boeckmann B, Capella-Gutierrez S, Dalquen DA, DeLuca T, Forslund K, Huerta-Cepas J, Linard B, Pereira C, Pryszcz LP, Schreiber F, da Silva AS, Szklarczyk D, Train CM, Bork P, Lecompte O, von Mering C, Xenarios I, Sjölander K, Jensen LJ, Martin MJ, Muffato M, Quest for Orthologs consortium. Gabaldón T, Lewis SE, Thomas PD, Sonnhammer E, Dessimoz C. Standardized benchmarking in the quest for orthologs. Nat Methods. 2016;13(5):425–430. doi: 10.1038/nmeth.3830. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
