

REX technologies for profiling and decoding the electrophile signaling axes mediated by Rosetta Stone proteins
- PMID: 32046846
- PMCID: PMC7027669
- DOI: 10.1016/bs.mie.2019.02.039
REX technologies for profiling and decoding the electrophile signaling axes mediated by Rosetta Stone proteins
Abstract
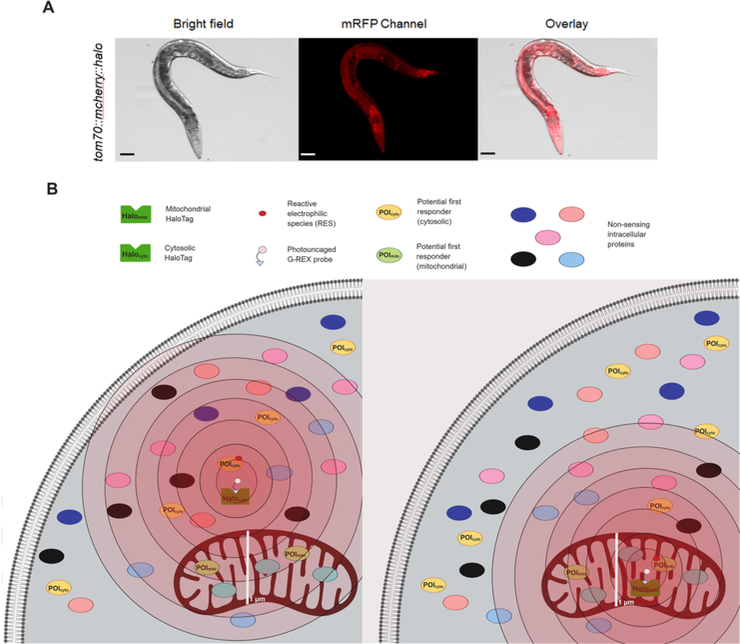
It is now clear that some cysteines on some proteins are highly tuned to react with electrophiles. Based on numerous studies, it is also established that electrophile sensing underpins rewiring of several critical signaling processes. These electrophile-sensing proteins, or privileged first responders (PFRs), are likely critically relevant for drug design. However, identifying PFRs remains a challenging and unsolved problem, despite the development of several high-throughput methods to ID proteins that react with electrophiles. More importantly, we remain unable to rank how different PFRs identified under different conditions relate to one another, in terms of sensing or signaling capacity. Here we evaluate different methods to assay sensing functions of proteins and discuss these methods in the context of developing a "ranking scheme." Based on theoretical and experimental evidence, we propose that T-REX-the only targeted-electrophile delivery tool presently available-is a reliable method to rank PFRs. Finally, we address to what extent electrophile sensing and downstream signaling are correlated. Based on our current data, we observe that such behaviors are indeed correlated. It is our hope that through this manuscript researchers from various arms of the stress signaling fields will focus on developing a quantitative understanding of precision electrophile labeling.
Keywords: Electrophile signaling; G-REX; Privileged first responders; Ranking reactivity; Redox-regulated pathways; Stress response; T-REX.
© 2020 Elsevier Inc. All rights reserved.
Figures






Similar articles
-
An Oculus to Profile and Probe Target Engagement In Vivo: How T-REX Was Born and Its Evolution into G-REX.Acc Chem Res. 2021 Feb 2;54(3):618-631. doi: 10.1021/acs.accounts.0c00537. Epub 2020 Nov 23. Acc Chem Res. 2021. PMID: 33228351
-
Getting the Right Grip? How Understanding Electrophile Selectivity Profiles Could Illuminate Our Understanding of Redox Signaling.Antioxid Redox Signal. 2020 Nov 20;33(15):1077-1091. doi: 10.1089/ars.2019.7894. Epub 2019 Nov 4. Antioxid Redox Signal. 2020. PMID: 31578876 Free PMC article.
-
Interrogating Precision Electrophile Signaling.Trends Biochem Sci. 2019 Apr;44(4):380-381. doi: 10.1016/j.tibs.2019.01.006. Epub 2019 Feb 11. Trends Biochem Sci. 2019. PMID: 30765181 Free PMC article.
-
Privileged Electrophile Sensors: A Resource for Covalent Drug Development.Cell Chem Biol. 2017 Jul 20;24(7):787-800. doi: 10.1016/j.chembiol.2017.05.023. Epub 2017 Jun 22. Cell Chem Biol. 2017. PMID: 28648380 Free PMC article. Review.
-
Z-REX: shepherding reactive electrophiles to specific proteins expressed tissue specifically or ubiquitously, and recording the resultant functional electrophile-induced redox responses in larval fish.Nat Protoc. 2023 May;18(5):1379-1415. doi: 10.1038/s41596-023-00809-8. Epub 2023 Apr 5. Nat Protoc. 2023. PMID: 37020146 Free PMC article. Review.
Cited by
-
Neighborhood watch: tools for defining locale-dependent subproteomes and their contextual signaling activities.RSC Chem Biol. 2020 May 27;1(2):42-55. doi: 10.1039/d0cb00041h. eCollection 2020 Jun 1. RSC Chem Biol. 2020. PMID: 34458747 Free PMC article. Review.
References
-
- Blatnik M, Frizzell N, Thorpe SR, Baynes JW, 2008. Inactivation of glyceraldehyde-3-phosphate dehydrogenase by fumarate in diabetes: Formation of S-(2-succinyl)cysteine, a novel chemical modification of protein and possible biomarker of mitochondrial stress. Diabetes 57 (1), 41–49. 10.2337/db07-0838. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
