

Basis for metabolite-dependent Cullin-RING ligase deneddylation by the COP9 signalosome
- PMID: 32047038
- PMCID: PMC7049131
- DOI: 10.1073/pnas.1911998117
Basis for metabolite-dependent Cullin-RING ligase deneddylation by the COP9 signalosome
Abstract
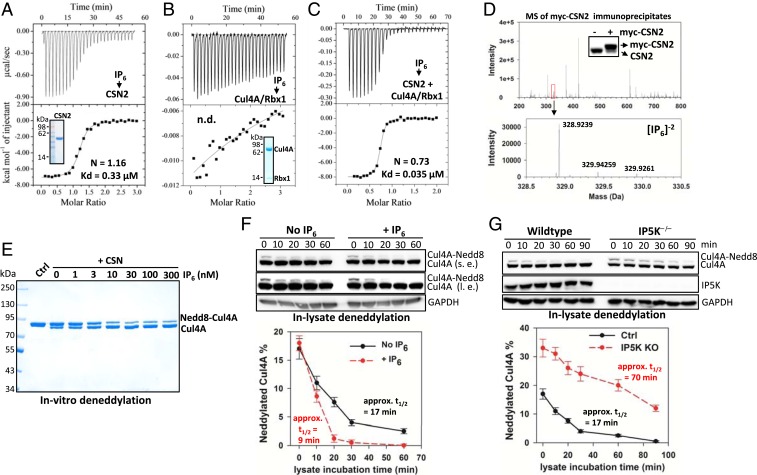
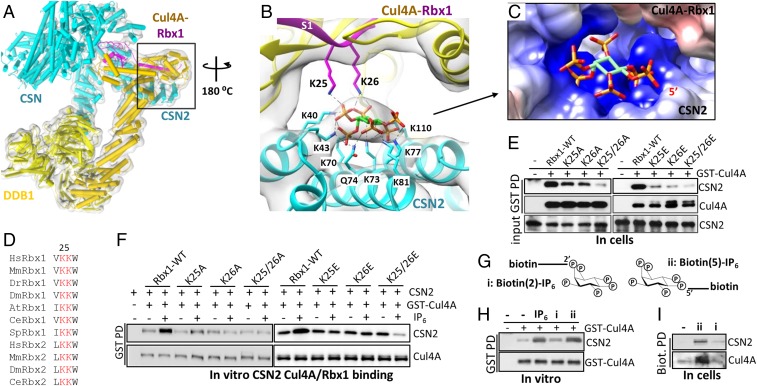
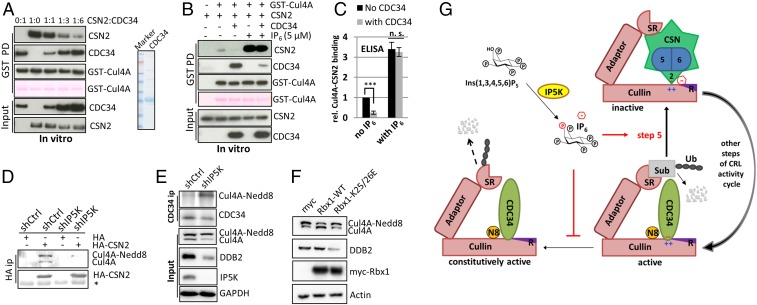
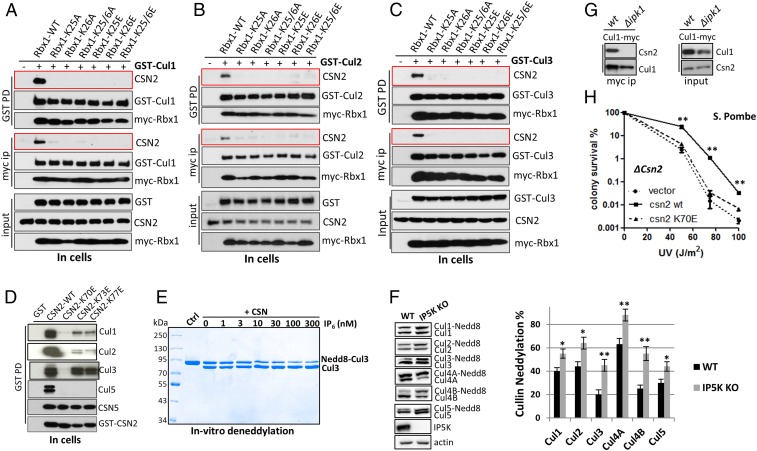
The Cullin-RING ligases (CRLs) are the largest family of ubiquitin E3s activated by neddylation and regulated by the deneddylase COP9 signalosome (CSN). The inositol polyphosphate metabolites promote the formation of CRL-CSN complexes, but with unclear mechanism of action. Here, we provide structural and genetic evidence supporting inositol hexakisphosphate (IP6) as a general CSN cofactor recruiting CRLs. We determined the crystal structure of IP6 in complex with CSN subunit 2 (CSN2), based on which we identified the IP6-corresponding electron density in the cryoelectron microscopy map of a CRL4A-CSN complex. IP6 binds to a cognate pocket formed by conserved lysine residues from CSN2 and Rbx1/Roc1, thereby strengthening CRL-CSN interactions to dislodge the E2 CDC34/UBE2R from CRL and to promote CRL deneddylation. IP6 binding-deficient Csn2K70E/K70E knockin mice are embryonic lethal. The same mutation disabled Schizosaccharomyces pombe Csn2 from rescuing UV-hypersensitivity of csn2-null yeast. These data suggest that CRL transition from the E2-bound active state to the CSN-bound sequestered state is critically assisted by an interfacial IP6 small molecule, whose metabolism may be coupled to CRL-CSN complex dynamics.
Keywords: COP9 signalosome; Cullin-RING ligases; deneddylation; inositol hexakisphosphate; intermolecular glue.
Conflict of interest statement
The authors declare no competing interest.
Figures

Comment in
-
Regulation of Cullin-RING E3 ligase dynamics by Inositol hexakisphosphate.Proc Natl Acad Sci U S A. 2020 Mar 24;117(12):6292-6294. doi: 10.1073/pnas.2001683117. Epub 2020 Mar 10. Proc Natl Acad Sci U S A. 2020. PMID: 32156730 Free PMC article. No abstract available.
Similar articles
-
IP6-assisted CSN-COP1 competition regulates a CRL4-ETV5 proteolytic checkpoint to safeguard glucose-induced insulin secretion.Nat Commun. 2021 Apr 28;12(1):2461. doi: 10.1038/s41467-021-22941-3. Nat Commun. 2021. PMID: 33911083 Free PMC article.
-
Are Inositol Polyphosphates the Missing Link in Dynamic Cullin RING Ligase Regulation by the COP9 Signalosome?Biomolecules. 2019 Aug 7;9(8):349. doi: 10.3390/biom9080349. Biomolecules. 2019. PMID: 31394817 Free PMC article. Review.
-
Cullin-RING Ligase Regulation by the COP9 Signalosome: Structural Mechanisms and New Physiologic Players.Adv Exp Med Biol. 2020;1217:47-60. doi: 10.1007/978-981-15-1025-0_4. Adv Exp Med Biol. 2020. PMID: 31898221 Review.
-
Inositol hexakisphosphate (IP6) generated by IP5K mediates cullin-COP9 signalosome interactions and CRL function.Proc Natl Acad Sci U S A. 2016 Mar 29;113(13):3503-8. doi: 10.1073/pnas.1525580113. Epub 2016 Mar 14. Proc Natl Acad Sci U S A. 2016. PMID: 26976604 Free PMC article.
-
The COP9 signalosome: A versatile regulatory hub of Cullin-RING ligases.Trends Biochem Sci. 2023 Jan;48(1):82-95. doi: 10.1016/j.tibs.2022.08.003. Epub 2022 Aug 27. Trends Biochem Sci. 2023. PMID: 36041947 Review.
Cited by
-
Suramin and NF449 are IP5K inhibitors that disrupt inositol hexakisphosphate-mediated regulation of cullin-RING ligase and sensitize cancer cells to MLN4924/pevonedistat.J Biol Chem. 2020 Jul 24;295(30):10281-10292. doi: 10.1074/jbc.RA120.014375. Epub 2020 Jun 3. J Biol Chem. 2020. PMID: 32493769 Free PMC article.
-
IP6-assisted CSN-COP1 competition regulates a CRL4-ETV5 proteolytic checkpoint to safeguard glucose-induced insulin secretion.Nat Commun. 2021 Apr 28;12(1):2461. doi: 10.1038/s41467-021-22941-3. Nat Commun. 2021. PMID: 33911083 Free PMC article.
-
Stable Isotopomers of myo-Inositol Uncover a Complex MINPP1-Dependent Inositol Phosphate Network.ACS Cent Sci. 2022 Dec 28;8(12):1683-1694. doi: 10.1021/acscentsci.2c01032. Epub 2022 Dec 5. ACS Cent Sci. 2022. PMID: 36589890 Free PMC article.
-
Dynamic molecular architecture and substrate recruitment of cullin3-RING E3 ligase CRL3KBTBD2.Nat Struct Mol Biol. 2024 Feb;31(2):336-350. doi: 10.1038/s41594-023-01182-6. Epub 2024 Feb 8. Nat Struct Mol Biol. 2024. PMID: 38332366 Free PMC article.
-
Inositol polyphosphate-protein interactions: Implications for microbial pathogenicity.Cell Microbiol. 2021 Jun;23(6):e13325. doi: 10.1111/cmi.13325. Epub 2021 Mar 25. Cell Microbiol. 2021. PMID: 33721399 Free PMC article. Review.
References
-
- Petroski M. D., Deshaies R. J., Function and regulation of cullin-RING ubiquitin ligases. Nat. Rev. Mol. Cell Biol. 6, 9–20 (2005). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
