

Nucleosome destabilization by nuclear non-coding RNAs
- PMID: 32047236
- PMCID: PMC7012929
- DOI: 10.1038/s42003-020-0784-9
Nucleosome destabilization by nuclear non-coding RNAs
Abstract
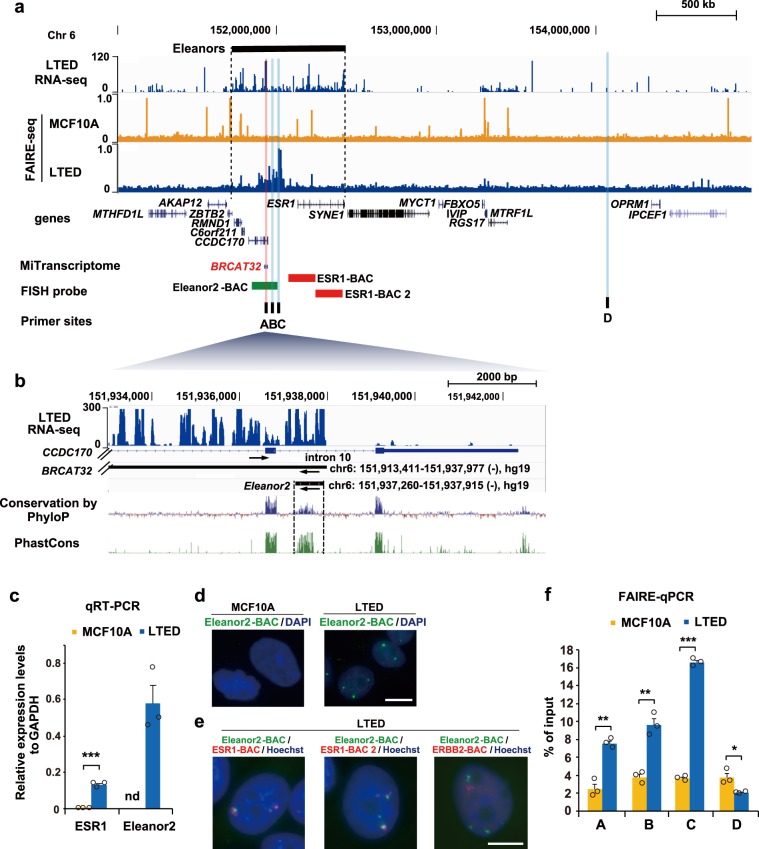
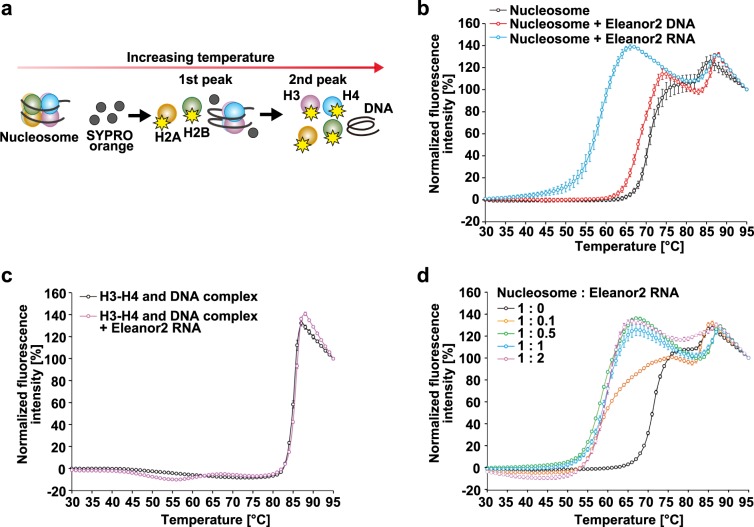
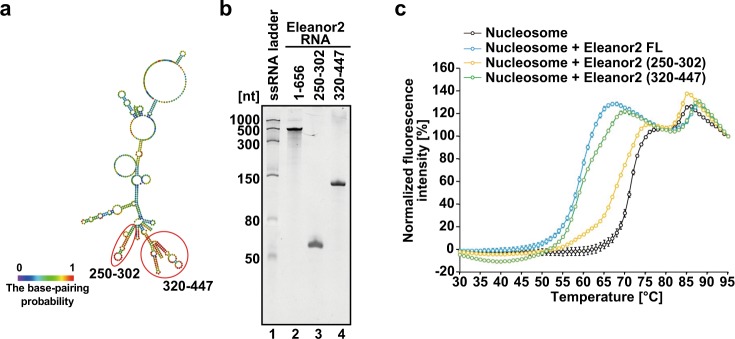
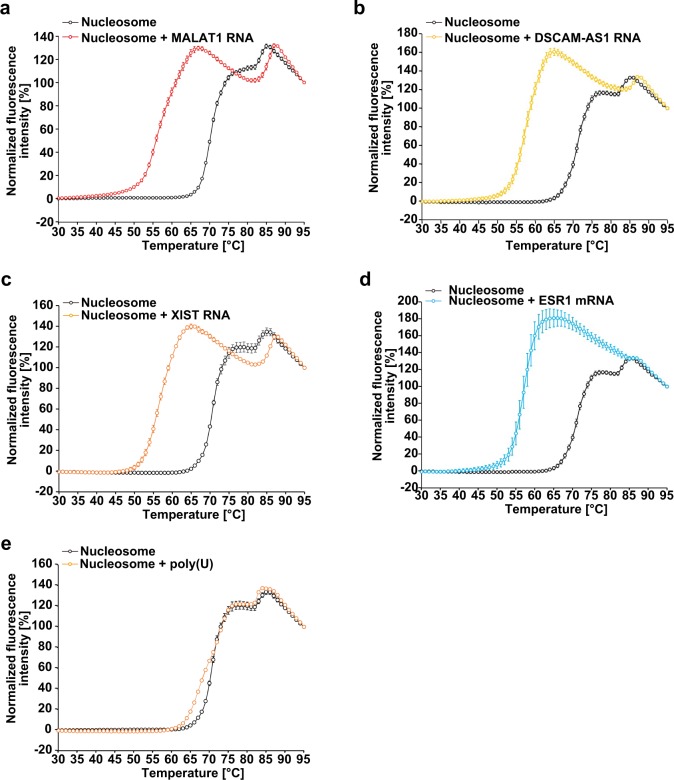
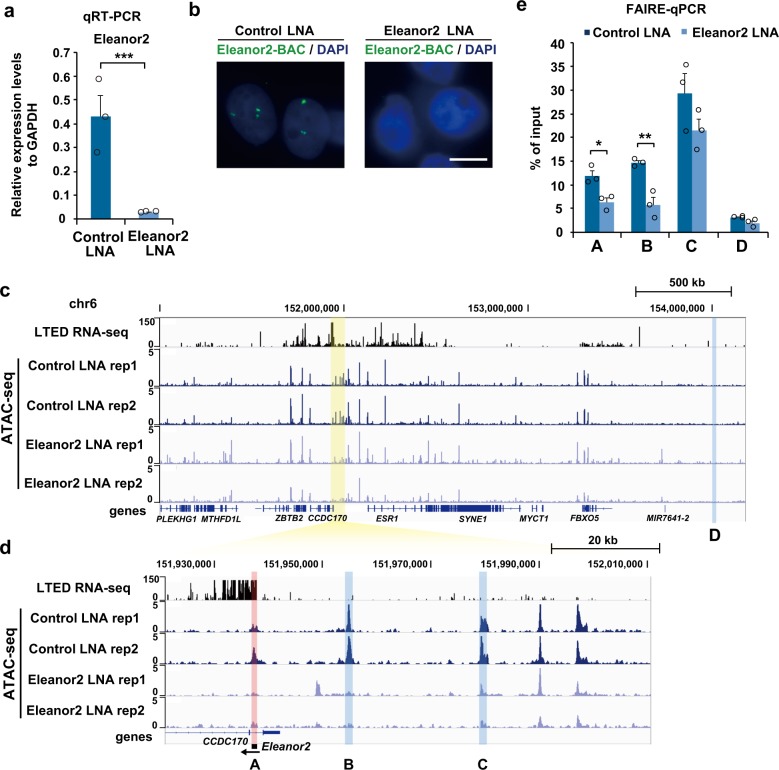
In the nucleus, genomic DNA is wrapped around histone octamers to form nucleosomes. In principle, nucleosomes are substantial barriers to transcriptional activities. Nuclear non-coding RNAs (ncRNAs) are proposed to function in chromatin conformation modulation and transcriptional regulation. However, it remains unclear how ncRNAs affect the nucleosome structure. Eleanors are clusters of ncRNAs that accumulate around the estrogen receptor-α (ESR1) gene locus in long-term estrogen deprivation (LTED) breast cancer cells, and markedly enhance the transcription of the ESR1 gene. Here we detected nucleosome depletion around the transcription site of Eleanor2, the most highly expressed Eleanor in the LTED cells. We found that the purified Eleanor2 RNA fragment drastically destabilized the nucleosome in vitro. This activity was also exerted by other ncRNAs, but not by poly(U) RNA or DNA. The RNA-mediated nucleosome destabilization may be a common feature among natural nuclear RNAs, and may function in transcription regulation in chromatin.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Wolffe AP. Chromatin: Structure & Function. London, UK: Academic Press; 1998.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
