

Transcriptional dysregulation in developing trigeminal sensory neurons in the LgDel mouse model of DiGeorge 22q11.2 deletion syndrome
- PMID: 32047912
- PMCID: PMC7158380
- DOI: 10.1093/hmg/ddaa024
Transcriptional dysregulation in developing trigeminal sensory neurons in the LgDel mouse model of DiGeorge 22q11.2 deletion syndrome
Erratum in
-
Transcriptional dysregulation in developing trigeminal sensory neurons in the LgDel mouse model of DiGeorge 22q11.2 deletion syndrome.Hum Mol Genet. 2020 Jun 3;29(9):1580. doi: 10.1093/hmg/ddaa067. Hum Mol Genet. 2020. PMID: 32358605 Free PMC article. No abstract available.
Abstract
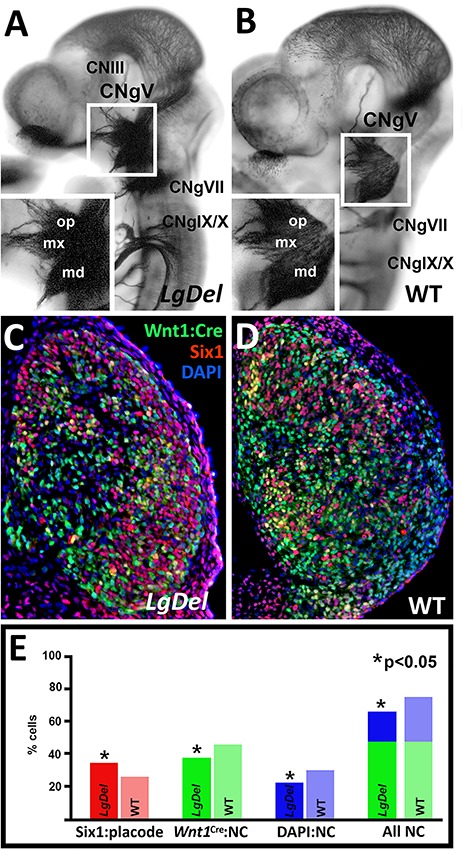
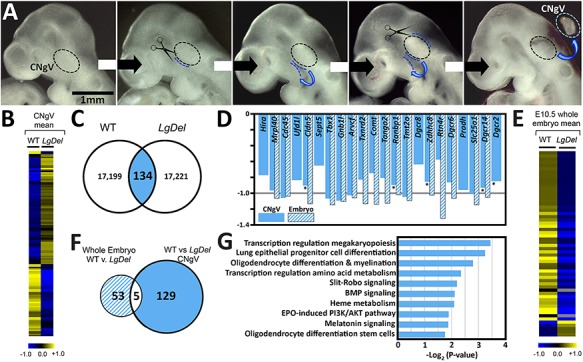
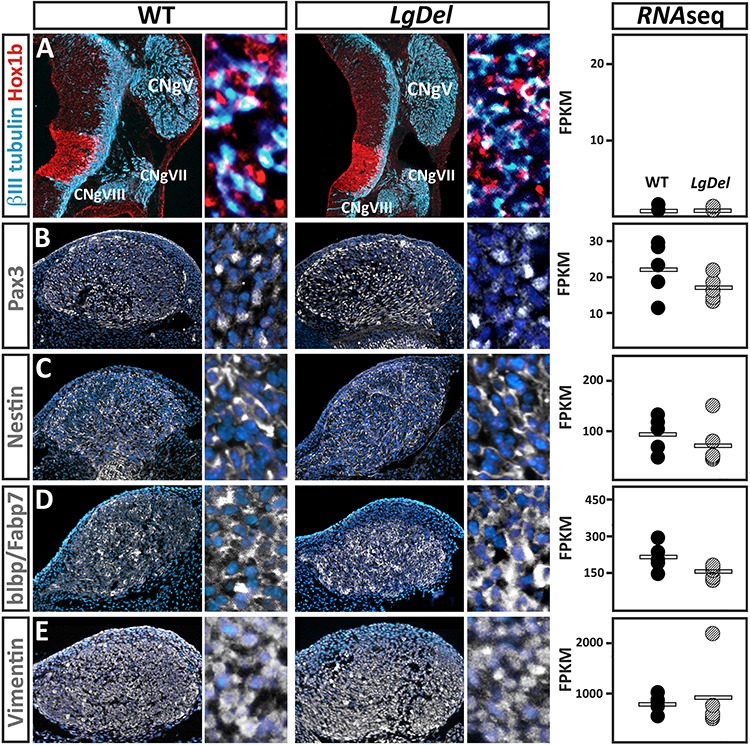
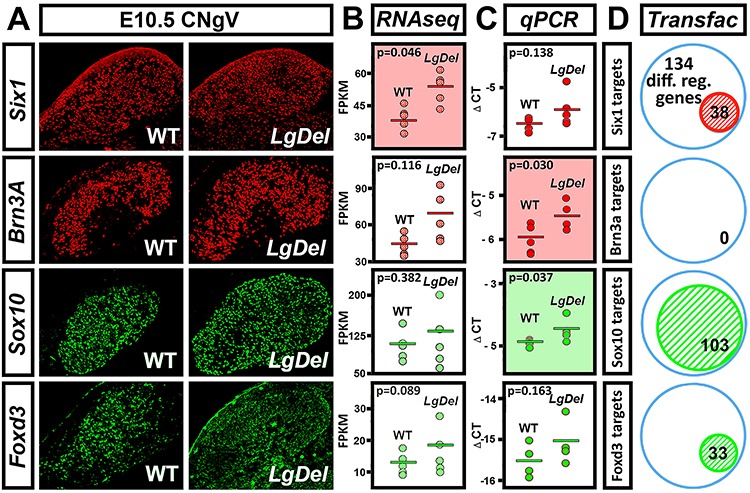
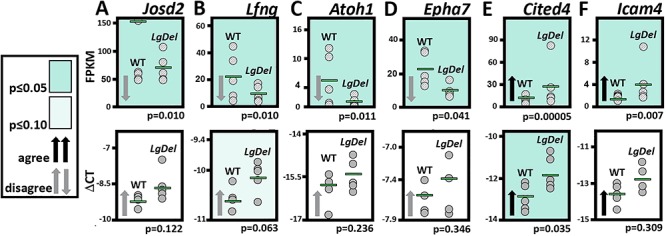
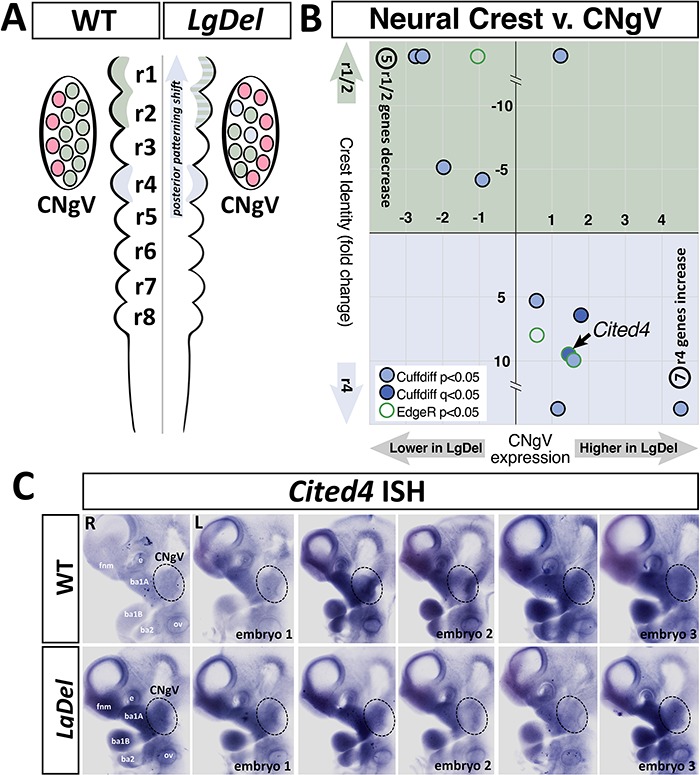
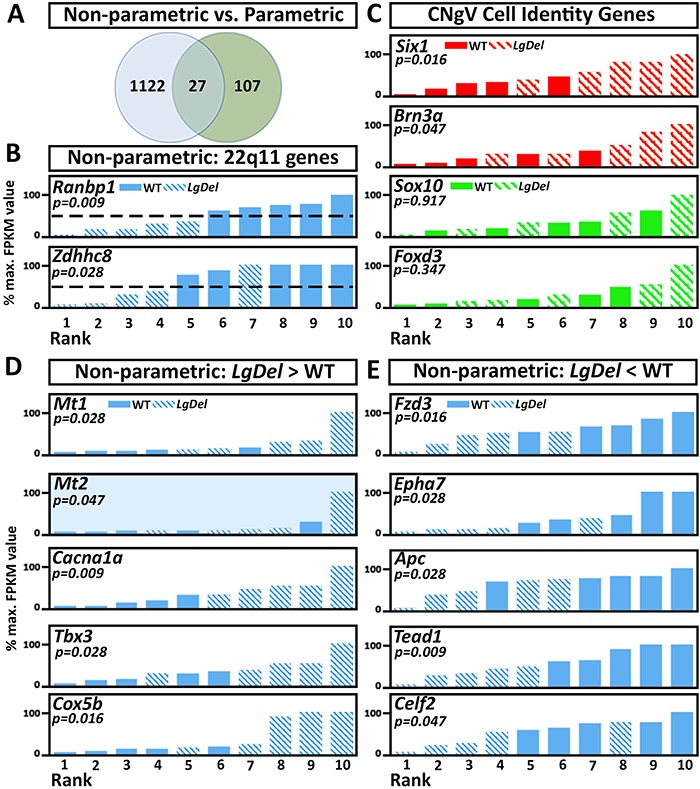
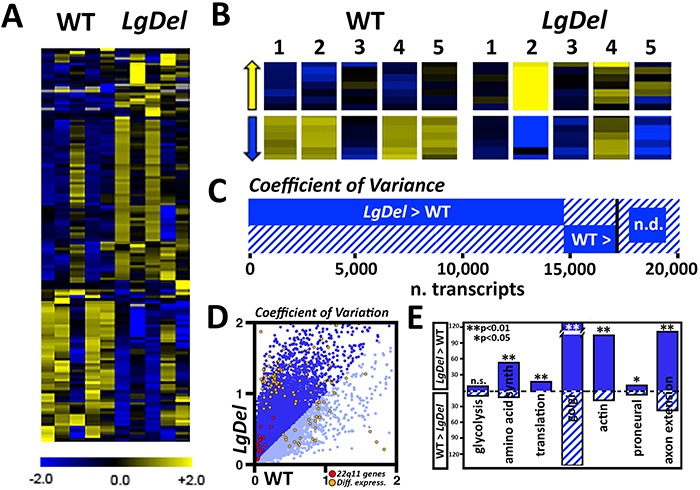
LgDel mice, which model the heterozygous deletion of genes at human chromosome 22q11.2 associated with DiGeorge/22q11.2 deletion syndrome (22q11DS), have cranial nerve and craniofacial dysfunction as well as disrupted suckling, feeding and swallowing, similar to key 22q11DS phenotypes. Divergent trigeminal nerve (CN V) differentiation and altered trigeminal ganglion (CNgV) cellular composition prefigure these disruptions in LgDel embryos. We therefore asked whether a distinct transcriptional state in a specific population of early differentiating LgDel cranial sensory neurons, those in CNgV, a major source of innervation for appropriate oropharyngeal function, underlies this departure from typical development. LgDel versus wild-type (WT) CNgV transcriptomes differ significantly at E10.5 just after the ganglion has coalesced. Some changes parallel altered proportions of cranial placode versus cranial neural crest-derived CNgV cells. Others are consistent with a shift in anterior-posterior patterning associated with divergent LgDel cranial nerve differentiation. The most robust quantitative distinction, however, is statistically verifiable increased variability of expression levels for most of the over 17 000 genes expressed in common in LgDel versus WT CNgV. Thus, quantitative expression changes of functionally relevant genes and increased stochastic variation across the entire CNgV transcriptome at the onset of CN V differentiation prefigure subsequent disruption of cranial nerve differentiation and oropharyngeal function in LgDel mice.
© The Author(s) 2020. Published by Oxford University Press.
Figures
Similar articles
-
Aberrant early growth of individual trigeminal sensory and motor axons in a series of mouse genetic models of 22q11.2 deletion syndrome.Hum Mol Genet. 2020 Nov 4;29(18):3081-3093. doi: 10.1093/hmg/ddaa199. Hum Mol Genet. 2020. PMID: 32901287 Free PMC article.
-
Selective disruption of trigeminal sensory neurogenesis and differentiation in a mouse model of 22q11.2 deletion syndrome.Dis Model Mech. 2022 Feb 1;15(2):dmm047357. doi: 10.1242/dmm.047357. Epub 2021 May 4. Dis Model Mech. 2022. PMID: 33722956 Free PMC article.
-
Dysphagia and disrupted cranial nerve development in a mouse model of DiGeorge (22q11) deletion syndrome.Dis Model Mech. 2014 Feb;7(2):245-57. doi: 10.1242/dmm.012484. Epub 2013 Dec 19. Dis Model Mech. 2014. PMID: 24357327 Free PMC article.
-
Three phases of DiGeorge/22q11 deletion syndrome pathogenesis during brain development: patterning, proliferation, and mitochondrial functions of 22q11 genes.Int J Dev Neurosci. 2011 May;29(3):283-94. doi: 10.1016/j.ijdevneu.2010.08.005. Epub 2010 Sep 15. Int J Dev Neurosci. 2011. PMID: 20833244 Free PMC article. Review.
-
Hard to swallow: Developmental biological insights into pediatric dysphagia.Dev Biol. 2016 Jan 15;409(2):329-42. doi: 10.1016/j.ydbio.2015.09.024. Epub 2015 Nov 7. Dev Biol. 2016. PMID: 26554723 Free PMC article. Review.
Cited by
-
Aberrant early growth of individual trigeminal sensory and motor axons in a series of mouse genetic models of 22q11.2 deletion syndrome.Hum Mol Genet. 2020 Nov 4;29(18):3081-3093. doi: 10.1093/hmg/ddaa199. Hum Mol Genet. 2020. PMID: 32901287 Free PMC article.
-
Advances in Swallowing Neurophysiology across Pediatric Development: Current Evidence and Insights.Curr Phys Med Rehabil Rep. 2021 Dec;9(4):267-276. doi: 10.1007/s40141-021-00334-3. Epub 2021 Nov 18. Curr Phys Med Rehabil Rep. 2021. PMID: 34956736 Free PMC article.
-
The 22q11.2 region regulates presynaptic gene-products linked to schizophrenia.Nat Commun. 2022 Jun 27;13(1):3690. doi: 10.1038/s41467-022-31436-8. Nat Commun. 2022. PMID: 35760976 Free PMC article.
-
Why Does the Face Predict the Brain? Neural Crest Induction, Craniofacial Morphogenesis, and Neural Circuit Development.Front Physiol. 2020 Dec 11;11:610970. doi: 10.3389/fphys.2020.610970. eCollection 2020. Front Physiol. 2020. PMID: 33362582 Free PMC article. Review.
-
Out of Line or Altered States? Neural Progenitors as a Target in a Polygenic Neurodevelopmental Disorder.Dev Neurosci. 2024;46(1):1-21. doi: 10.1159/000530898. Epub 2023 May 10. Dev Neurosci. 2024. PMID: 37231803 Free PMC article. Review.
References
-
- Cooper-Brown L., Copeland S., Dailey S., Downey D., Petersen M.C., Stimson C. and Van Dyke D.C. (2008) Feeding and swallowing dysfunction in genetic syndromes. Dev. Disabil. Res. Rev., 14, 147–157. - PubMed
-
- Kakodkar K. and Schroeder J.W. Jr. (2013) Pediatric dysphagia. Pediatr. Clin. North Am., 60, 969–977. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
