

NAD+ Repletion Rescues Female Fertility during Reproductive Aging
- PMID: 32049001
- PMCID: PMC7063679
- DOI: 10.1016/j.celrep.2020.01.058
NAD+ Repletion Rescues Female Fertility during Reproductive Aging
Abstract
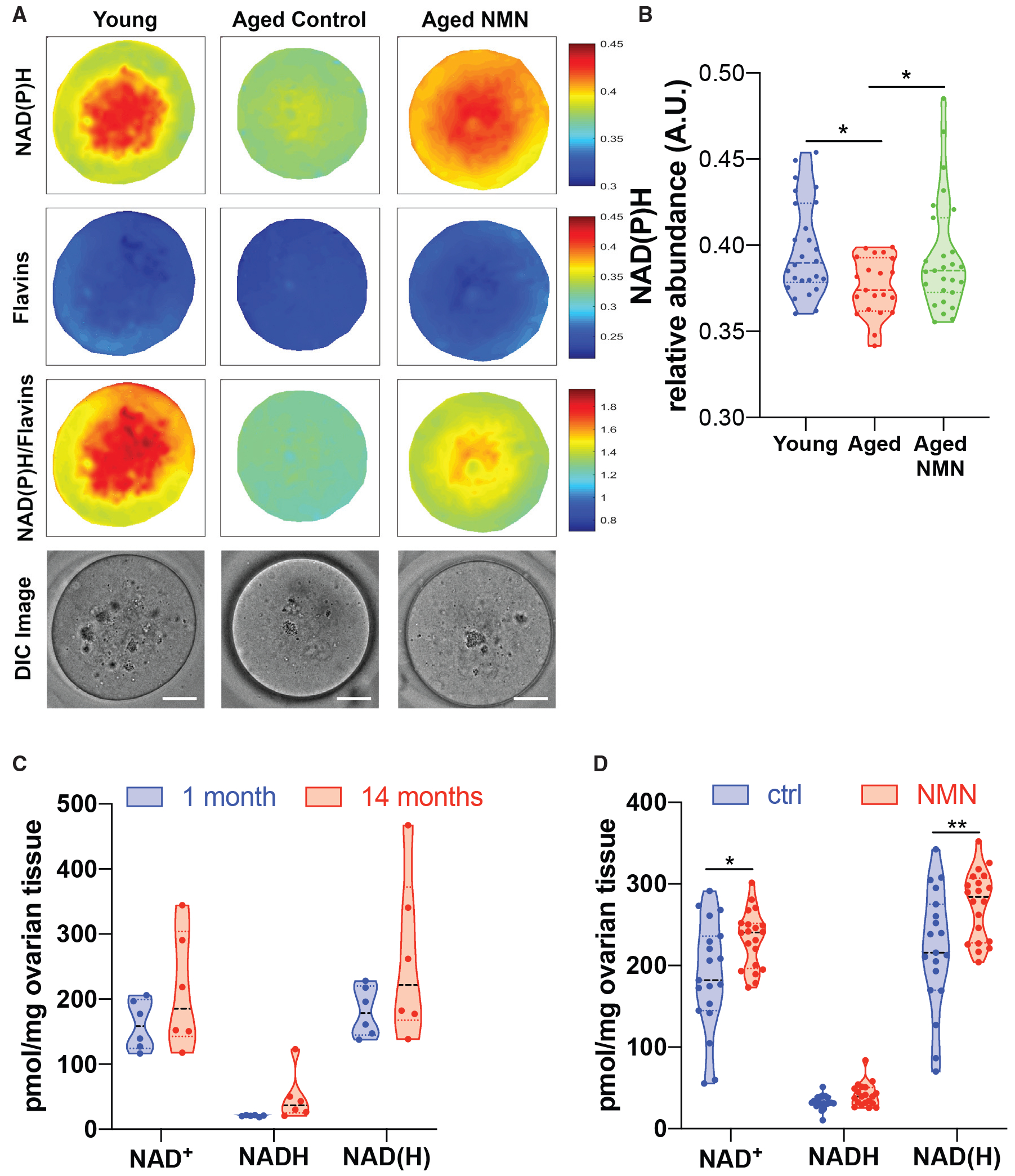
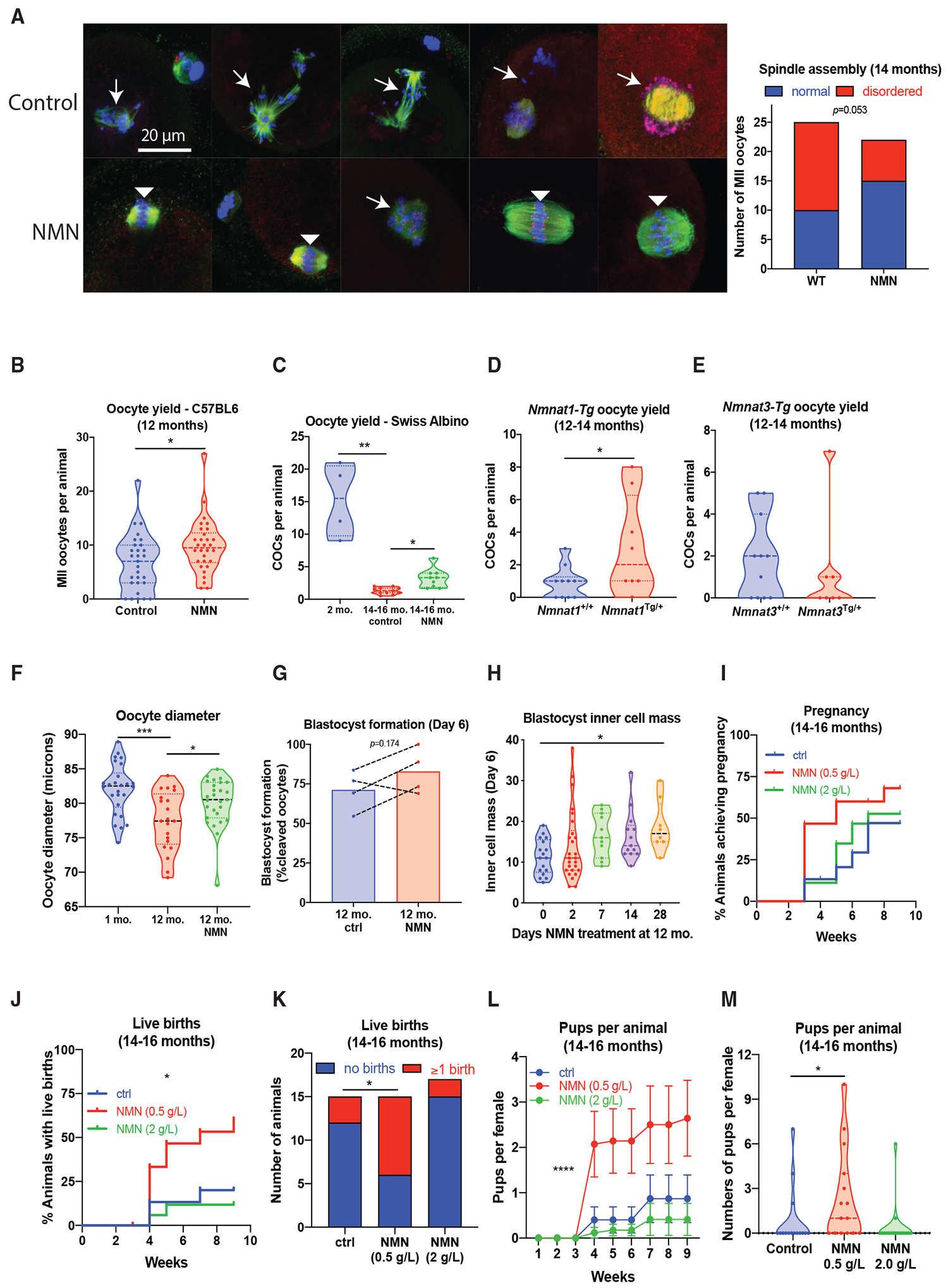
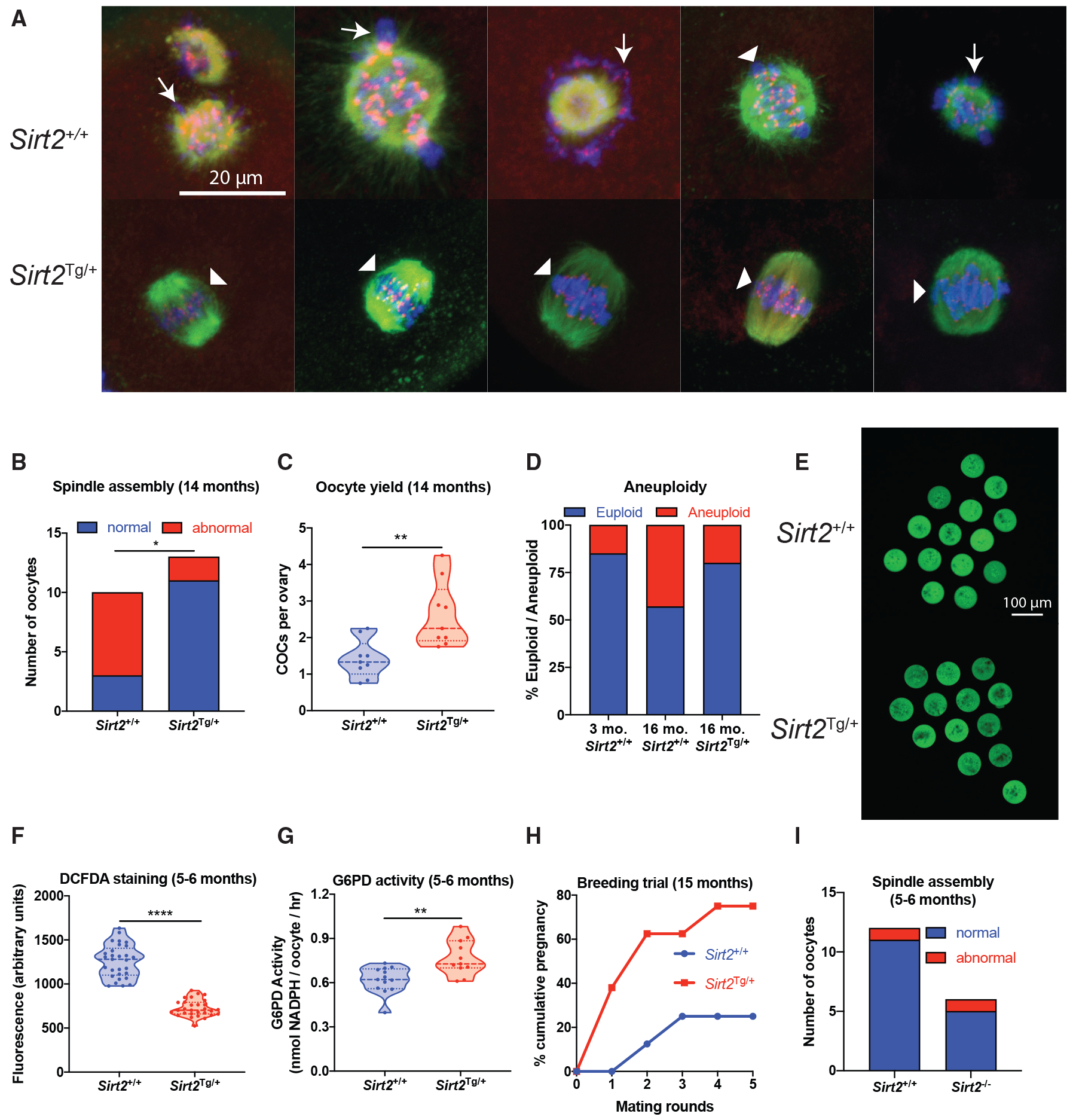
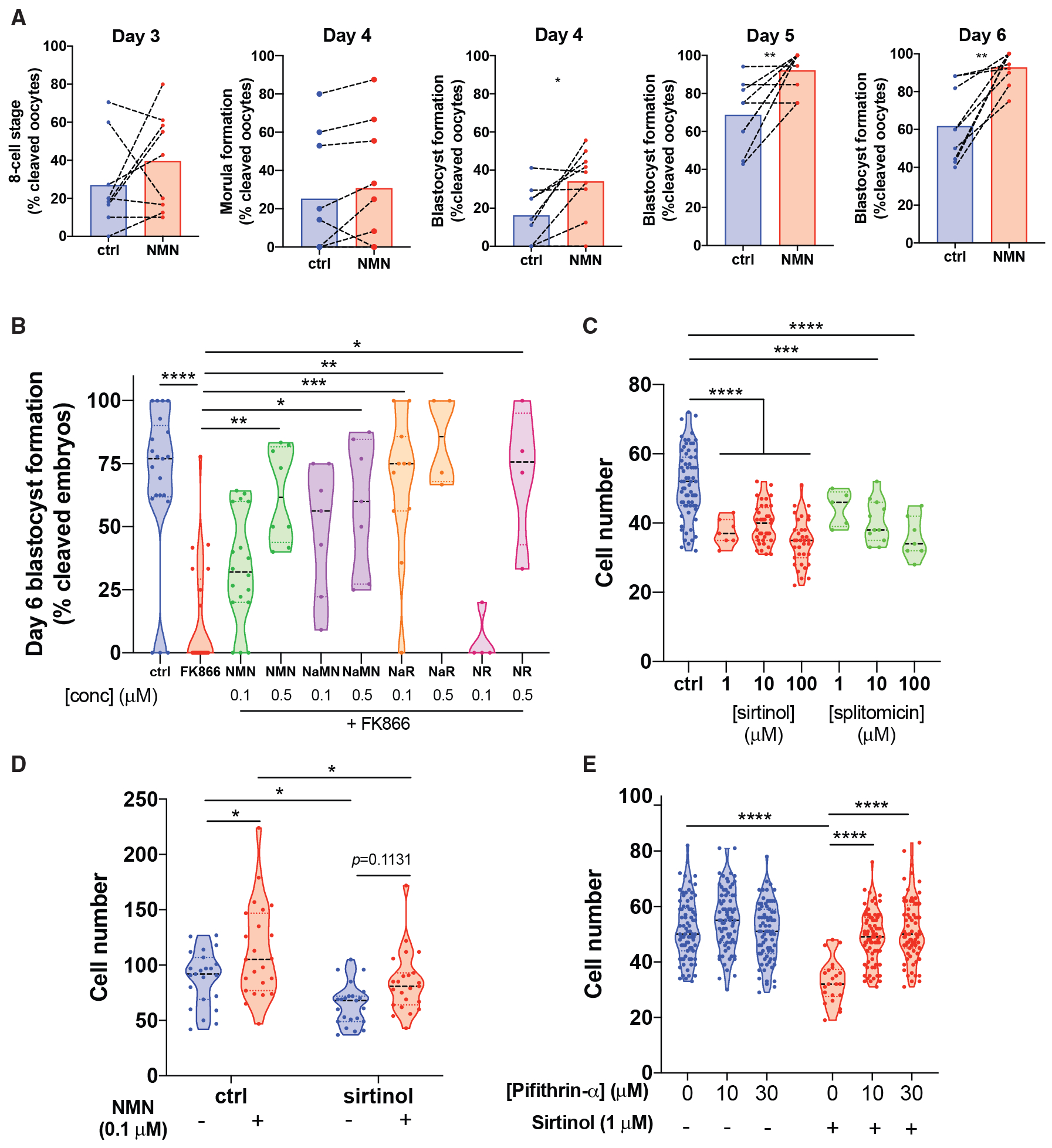
Reproductive aging in female mammals is an irreversible process associated with declining oocyte quality, which is the rate-limiting factor to fertility. Here, we show that this loss of oocyte quality with age accompanies declining levels of the prominent metabolic cofactor nicotinamide adenine dinucleotide (NAD+). Treatment with the NAD+ metabolic precursor nicotinamide mononucleotide (NMN) rejuvenates oocyte quality in aged animals, leading to restoration in fertility, and this can be recapitulated by transgenic overexpression of the NAD+-dependent deacylase SIRT2, though deletion of this enzyme does not impair oocyte quality. These benefits of NMN extend to the developing embryo, where supplementation reverses the adverse effect of maternal age on developmental milestones. These findings suggest that late-life restoration of NAD+ levels represents an opportunity to rescue female reproductive function in mammals.
Keywords: SIRT2; aging; embryo development; female fertility; in vitro fertilization; infertility; nicotinamide adenine dinucleotide (NAD+); nicotinamide mononucleotide (NMN); oocyte; reproductive aging.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests L.E.W., H.A.H., and D.A.S. are cofounders, shareholders, directors, and advisers of Jumpstart Fertility Inc., which was founded to develop the work described here, and are inventors on a patent (WO2019023748A1 “Methods to improve fertility”) that forms the basis of this work and has been licensed to Jumpstart Fertility. D.A.S. is an inventor on a patent (WO2013002880A1) that was previously licensed to Jumpstart Fertility. The salaries of M.J.B. and D.M.G. working in the labs of L.E.W. and R.B.G. at UNSW were paid by sponsored research from Jumpstart Fertility to UNSW. K.S. was an employee of Jumpstart Fertility. W.-H.J.H. and D.R.L. hold shares in Jumpstart Fertility. H.A.H. undertakes ART in the private sector in affiliation with Queensland Fertility Group. L.E.W. and D.A.S. are also advisers and shareholders in EdenRoc Sciences (Metro Biotech NSW, Metro Biotech, Liberty Biosecurity) and in Life Biosciences LLC and its daughter companies (Jumpstart Fertility, Continuum Biosciences, Senolytic Therapeutics, Selphagy, Animal Biosciences, Iduna). L.E.W. provides consulting work for Life Biosciences and is an adviser and shareholder in Intravital Pty Ltd. D.A.S. is also a founder, equity owner, adviser, director, consultant, investor, and/or inventor on patents licensed to Vium, Jupiter Orphan Therapeutics, Cohbar, Galilei Biosciences, Wellomics, EdenRoc Sciences (and affiliates Arc-Bio, Dovetail Genomics, Claret, Revere Biosciences, UpRNA, MetroBiotech, Liberty Biosecurity), Life Biosciences (and affiliates Selphagy, Senolytic Therapeutics, Spotlight Therapeutics, Animal Biosciences, Iduna, Continuum, Jumpstart Fertility, Iduna). D.A.S. is an inventor on a patent application filed by Mayo Clinic and Harvard Medical School that has been licensed to Elysium Health. For details see https://genetics.med.harvard.edu/sinclair.
Figures
References
-
- Berger F, Lau C, Dahlmann M, and Ziegler M (2005). Subcellular compartmentation and differential catalytic properties of the three human nicotinamide mononucleotide adenylyltransferase isoforms. J. Biol. Chem 280, 36334–36341. - PubMed
-
- Bieganowski P, and Brenner C (2004). Discoveries of nicotinamide riboside as a nutrient and conserved NRK genes establish a Preiss-Handler independent route to NAD+ in fungi and humans. Cell 117, 495–502. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
